

")


Intramiocardial administration of resident c-kit+ cardiac progenital cells activates epicardial progenitor cells and promotes myocardial vascularation after the infarction
- 作者: Dergilev K.V1, Tsokolaeva Z.I1, Beloglazova I.B1,2, Zubkova E.S1,2, Boldyreva M.A1,2, Ratner E.I1, Diykanov D.T2, Menshikov M.U1, Parfenova E.V1,2
-
隶属关系:
- Russian Cardiology Research and Production Complex
- M.V. Lomonosov Moscow State University
- 期: 卷 13, 编号 1 (2018)
- 页面: 75-81
- 栏目: Articles
- ##submission.dateSubmitted##: 05.01.2023
- ##submission.datePublished##: 15.03.2018
- URL: https://genescells.ru/2313-1829/article/view/120736
- DOI: https://doi.org/10.23868/201805009
- ID: 120736

如何引用文章
全文:

详细
全文:
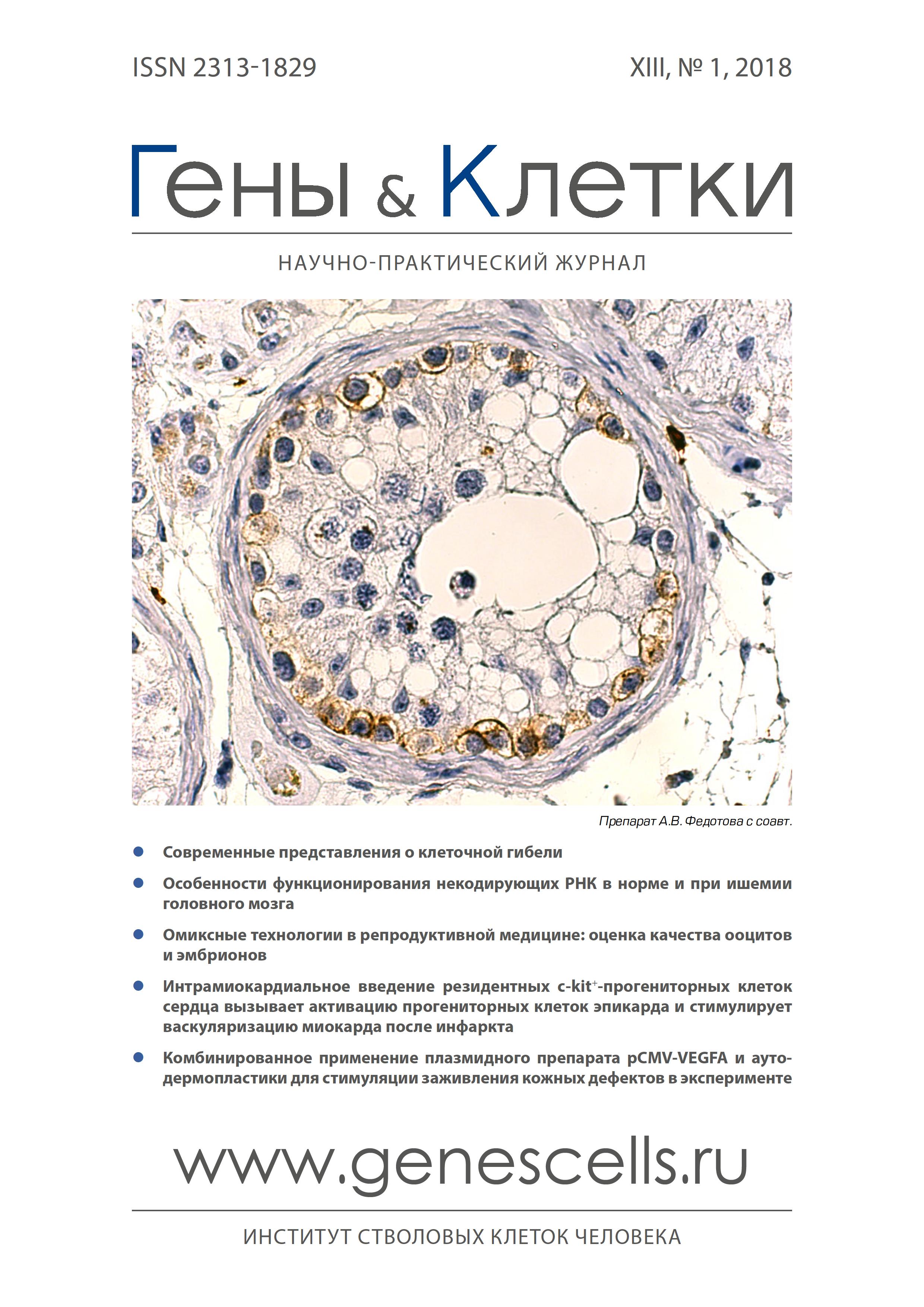
Введение сердечная недостаточность, возникающая в большинстве случаев после инфаркта миокарда (ИМ), продолжает лидировать среди основных причин смерти и инва-лидизации во всем мире [1]. Несмотря на повсеместное внедрение подходов для лечения острой коронарной окклюзии, связанных со стентированием или хирургической васкуляризацией, эти методы применимы только при условии наличия «подходящих» для вмешательства коронарных сосудов и сохранения жизнеспособности миокарда. В случае необратимого повреждения миокарда, восстановить кровоснабжение миокарда с помощью Гены & Клетки Том XIII, № 1, 2018 76 ОРИГИНАЛЬНЫЕ ИССЛЕДОВАНИЯ вышеуказанных методов не представляется возможным, что приводит к ремоделированию сердца и развитию сердечной недостаточности [2]. Образование новых кровеносных сосудов является частью естественного процесса регенерации миокарда, регулирует процесс постинфарктного ремоделирования и определяет прогноз для пациентов после перенесенного им [3, 4]. на сегодняшний день были исследованы различные подходы к стимуляции васкуляризации по-стинфарктного сердца, включая генную терапию, а также локальную/системную доставку ангиогенных факторов роста, результаты которых, к сожалению, оказались весьма спорными и имеющими ограниченную эффективность [5-7]. В связи с этим внимание исследователей было сосредоточено на поиске стволовых клеток, которые могут дифференцироваться в клетки сосудов и стимулировать васкуляризацию за счет паракринных эффектов, что может способствовать формированию сосудистой сети de novo, обеспечивающей выживаемость ишемизированных/гибернированных кардиоми-оцитов, поддерживать систолическую функцию сердца и предотвращать развитие сердечной недостаточности. Определенные надежды в этой области возлагаются на резидентные прогениторные клетки сердца (ПКС), которые являются активными участниками регенеративных процессов в миокарде [8, 9], как за счет индукции дифференцировки в основные клеточные типы миокарда, прежде всего в клетки сосудов, так и за счет паракрин-ных эффектов, включающих секрецию внеклеточных везикул [10, 11]. В экспериментальных и клинических работах продемонстрирована возможность ПКС существенно улучшать функцию сердца и стимулировать его васкуляризацию [l0, 11]. Однако, механизмы участия ПКС в регенерации миокарда, и в частности, в его неова-скуляризации после инфаркта изучены недостаточно. Цель исследования - оценить влияние интрамиокар-диальной трансплантации ПКС на активацию васкуло-генного пула клеток эпикарда. Материал и методы Экспериментальные животные Эксперименты проводили на самцах крыс линии Wistar (n=30), приобретенных в питомнике лабораторных животных «Пущино» (Пущино, Россия). Животные содержались в условиях вивария при свободном доступе к пище и воде в соответствии с требованиями ГОСТ. Эвтаназия крыс проводилась после ингаляционной наркотизации изофлюраном (Piramal Healthcare, Индия) методом дислокации шейного отдела позвоночника. Все необходимые манипуляции выполняли в соответствии с Директивой ЕС 2010/63/ЕС по экспериментам на животных и были одобрены этическим комитетом института (Национальный центр медицинских исследований кардиологии, разрешение № 385.06.2009). Получение и культивирование резидентных ПКС С-№-ПКС крысы были получены с использованием модифицированного метода, описанного ранее [12] ПКС выделяли из крыс-самцов линии Wistar (250-300 г). Животных наркотизировали ингаляцией изофлюраном, сердца извлекали, промывали в стерильном растворе фосфатно-солевого буфера (ФСБ), измельчали ножницами до кусочков размером 2-3 мм3 и инкубировали в течение 15 мин. в смеси 0,1 % коллагеназы A (Roche Diagnostics, США) и 0,2 % трипсина (Invitrogen, США). Обработанные образцы миокарда культивировали в условиях эксплантной культуры в течение 10 сут. с использованием питательной среды DMEM/F12 (Gibco, США), содержащей 10 % ФБС (ATCC, США), 10 нг/мл LIF (Sigma, США), 100 ЕД/мл пенициллина (Gibco, США), 100 ЕД/ мл стрептомицина (Gibco, США), 2 мМ L-глутамина (Gibco, США). С-№-ПКС были выделены из клеток эксплантной культуры с помощью метода иммуномагнитной селекции с использованием набора для выделения клеток сердца и протокола фирмы Miltenyi Biotec (США). На первом этапе была проведена деплеция клеток костного мозга с помощью антител к CD45 (BD, США) и антител, конъюгированных с магнитной бусиной (Miltenyi Biotec, США). Затем CD45(-) клетки сортировали с использованием антител против c-kit (Santa Cruz, США) и вторичных антител, конъюгированных с магнитной бусиной (Miltenyi Biotec, США). Изолированные клетки культивировали в чашках Петри, покрытых фибронектином, в среде DMEM/F12, содержащей 10 % ФБС, 100 ЕД/мл пенициллина, 100 ЕД/ мл стрептомицина, 2 мМ L-глутамина, 2 % B27 (Sigma, США), 1х инсулин-трансферрин-селинат (Gibco, США) и факторы роста: 20 нг/мл bFGF (Peprotech, США), 20 нг/ мл EGF (Peprotech, США), 10 нг/мл LIF (Sigma, США). Анализ иммунофенотипа ПКС методом проточной цитофлуориметрии Для проверки чистоты клеточной культуры и иммунофенотипа ПКС использовали цитофлуориметр FACS Canto II (BD, США). Для проведения анализа клетки центрифугировали (200 g, 5 мин.), супернатант удаляли, осадок инкубировали в течение 30 мин. при +4 °С в растворе 1 % BSA/ФСБ с антителами к одному из следующих антигенов: c-kit (Santa Cruz, США), CD31 (BD, США), CD34 (Santa Cruz, США) или CD45 (BD, США). После инкубации клетки центрифугировали (200 g, 5 мин.), дважды промывали ФСБ и ресуспендировали в 1 % BSA/ ФСБ с вторичными антителами, конъюгированными с AlexaFluor488 (Invitrogen, США). После 30-мин. инкубации при +4 °С клетки промывали, ресуспендировали в ФСБ и исследовали с помощью FACS Canto II (BD, США). Анализ иммунофенотипа ПКС методом иммуноцитохимии ПКС, культивированные на стеклах, покрытых фибронектином, фиксировали 3,7 % параформальдегидом, обрабатывали 0,1 % Triton X-100 в ФСБ и инкубировали с кроличьими поликлональными антителами к маркерам c-kit (Santa Cruz, США, 1:100, 2 ч. при 37 °C), Oct 3/4 (Abcam, США, 1: 100, 2 ч. при 37 °C), Nanog (Santa Cruz, США, 1:100, 2 ч. при 37 °С), а затем с вторичными антителами Alexa Fluor 488/594 (кат # A11001, A21207, Invitrogen, США, 1:800, 1 ч. при 37 °C). Ядра клеток были окрашены DAPI. Анализ дифференцировочной способности ПКС С-№-ПКС 3 пассажа высевали на культуральные чашки Петри, покрытые ламинином, с плотностью 20х103 кл/ см2 в среду a-MEM (Gibco, США), содержащую 10 % ФБС, 10 мкМ 5-азацитидина (Sigma, США) и инкубировали в течение 72 ч. с ежедневной сменой среды. Затем клетки культивировали в среде a-MEM (Gibco, США) с 10 % ФБС и 10 нг/мл TGF-ß (R&D, США) в течение 21 сут. со сменой среды каждые 72 ч. Дифференцировку С-№-ПКС анализировали иммуноокрашиванием антителами к тяжелым цепям миозина (Abcam, США, 1:100, 1 ч. при 37 °C) и фаллоидину-Alexa 488 (Invitrogen, США, 1: 100, 1 ч. при 37 °C), и вторичными антителами Alexa Fluor 488 (Invitrogen, США, 1:800, 1 ч. при 37 °C). Для индукции эндотелиальной дифференцировки С-№-ПКС 3 пассажа высевали на культуральные чашки Петри в среду DMEM/F12, содержащую 1% ФБС, Гены & Клетки Том XIII, № 1, 2018 ОрИГИНАльНыЕ ИсслЕдОВАНИЯ 77 1 % пенициллина/стрептомицина, 2 мМ L-глутамина, 10-8 M дексаметазона, 10 нг/мл фактора роста VEGF и культивировали 21 сут. с заменой дифференцировоч-ной среды каждые 48 ч. Эндотелиальную дифферен-цировку тестировали путем окрашивания клеток антителами к маркеру Pecam (CD31) (BD, сША, 1:100, 1 ч. при 37 °C) и вторичными антителами Alexa Fluor 488 (Invitrogen, сША, 1:800, 1 ч. при 37 °с). Для индукции гладкомышечной дифференцировки с-№-ПКс 3 пассажа высевали на культуральные чашки Петри в среду DMEM/F12, содержащую 1°% ФБс, 1 % пенициллина/стрептомицина, 2 мМ L-глутамина, 10-8 М дексаметазона и 40 нг/мл фактора роста PDGF BB (R&D, сША) и культивировали в течение 21 сут. с заменой дифференцировочной среды каждые 48 ч. Гладкомышечную дифференцировку тестировали окрашиванием клеток антителами к маркеру Gata 6 (Santa Cruz, сША, 1: 100, 1 ч. при 37 °C), гладкомышечному актину-FITC (Sigma-Aldrich, сША, 1:100 , 1 ч. при 37 °с) и вторичными антителами Alexa Fluor 488 (Invitrogen, сША, 1:800, 1 ч. при 37 °C). Моделирование инфаркта миокарда и трансплантация клеток Инфаркт миокарда был индуцирован перевязкой передней нисходящей коронарной артерии крыс Wistar по методу, описанному ранее [13]. Перед трансплантацией ПКс метили флуоресцентным красителем CM Dil (Invitrogen, сША) в соответствии с рекомендациями фирмы-изготовителя реагента. Клеточный препарат ПКс трансплантировали с помощью ин-трамиокардиального способа введения (4 инъекции по 25х104 кл/50 мкл среды M199 (Gibco, сША) после перевязки передней нисходящей коронарной артерии у взрослых крыс (опытная группа, n=15). Животным контрольной группы (n=15) интрамиокардиально выполняли 4 инъекции (по 50 мкл каждая) среды M199 без добавок. Животных выводили из эксперимента через 14 сут. после трансплантации клеточного препарата. Морфометрический анализ Перед забором сердец в полость миокарда ЛЖ вводили 0,1 мл насыщенного раствора KCl, что приводило к остановке сокращений в диастолу. Предсердия и крупные сосуды иссекали, сердца промывали нормальным физиологическим раствором, помещали в криосреду Tissue-Tek O.C.T. Compound (Sakura Finetek USA Inc, Япония) и замораживали в жидком азоте. Криосрезы сердец (толщина среза 7 мкм, интервал между срезами в поперечной плоскости от верхушки до основания левого желудочка - 300 мкм) хранили при -700 с. Криосрезы окрашивали с помощью метода Мэллори с использованием следующих растворов: раствор A (1 % кислый фуксин), B (1 % фосфомолибденовая кислота) и C (2 % оранжевый G, 0,5 % метиловый синий, 2 % щавелевая кислота). Зафиксированные криосрезы инкубировали последовательно в растворе А (2 мин.), растворе В (4 мин.) и растворе с (15 мин.), промывая дистиллированной водой после каждого красителями, обезвоживали и монтировали, используя среду на основе ксилола. Количественный анализ размеров рубца и других морфометрических параметров выполняли с помощью пакета программ MetaMorph® Microscopy Automation & Image Analysis Software (сША). Размер инфаркта оценивали на основании подсчета площади рубца по отношению к площади левого желудочка, выраженного в процентах. Для подсчета средней толщины поврежденной стенки ЛЖ использовали измерение толщины рубца в 3 местах. Полученные значения усреднялись. Для количественного определения степени дилатации ЛЖ применяли модифицированный метод Хохмана [14]: Индекс дилатации = (площадь полости ЛЖ/общая площадь ЛЖ) х (средняя толщина неповрежденной области ЛЖ/средняя толщина стенки ЛЖ в зонах риска). Индекс трансмурального поражения рассчитывали по формуле [14]: Индекс трансмурального поражения (%) = длина трансмурального инфаркта, затрагивающая эпикард/ общая длина инфаркта. Гистологическая оценка состояния трансплантированных клеток, васкуляризации и активации клеток эпикарда Замороженные криосрезы миокарда фиксировали 3,7 % параформальдегидом (20 мин.) и промывали ФсБ (5 мин.). При проведении оценки внутриклеточных маркеров дополнительно проводилась обработка срезов 0,1% раствором Тритона X-100 (5 мин.). срезы миокарда блокировали раствором, содержащим 1 % БсА, 10 % сыворотки донора вторых антител в ФсБ (30 мин.). После этого криосрезы окрашивали антителами к маркерам Ki-67 (Abcam, сША), cleaved caspase-3 (Cell signaling, сША), Pecam (CD31) (Becton Dickinson, сША), a-SMA FITC (Sigma-Aldrich, сША) или WT1 (Abcam, сША) в течение 1 ч., промывали и окрашивали антителами, конъюгированными с AlexaFluor488 (Invitrogen, сША, 1:800, 1 ч. при 37 °C). Анализ плотности капиллярной сети в периинфаркт-ной области включал подсчет CD31-позитивных структур на мм2 площади периинфарктной области; количество артериолов на мм2 оценивалось как количество a-SMA-положительных сосудов с хорошо видимым CD31 + внутренним слоем. Подсчет проводили в программе Imaje J software. Количество Wt1 + клеток эпикарда оценивали на срезах миокарда, полученных на 14 сут. после трансплантации ПКс или среды M199, с помощью ручного подсчета на мм2 площади эпикарда в программе Imaje J software. Полимеразная цепная реакция в реальном времени Рибонуклеиновую кислоту выделяли из с-№+-ПКс 3 пассажа с помощью набора для выделения РНК Quiagen (Quiagen, сША). синтез комплиментарной дезок-сирибонулеиновой кислоты проводили с применением набора реагентов «Maxima First Strand cDNA Synthesis Kit» (ThermoScientific, сША). Полимеразную цепную реакцию осуществляли на амплификаторе StepOnePlus Real-Time PCR System (ThermoScientific, сША) по стандартному протоколу с использованием следующих праймеров: HGF (for: CCAGCTAGAAACAAAGACTTGAAAGA rev: GAAATGTTTAAGATCTGTTTGCGTT), VEGF (for: ACTG-GACCCTGGCTTTACTG; rev: ACGCACTCCAGGGCTTCATC), ангиопоэтин-1 (for: GTGGCTGGAAAAACTTGAGA rev: TGGATTTCAAGACGGGATGT) и ангиопоэтин-2 (for: GAC-CAGTGGGCATCGCTACG rev: CTGGTTGGCTGATGCTACTG). Микроскопия и анализ изображений Клетки и криосрезы миокарда анализировали на флуоресцентном микроскопе Zeiss Axiovert 200 M (Carl Zeiss, Германия) с программным обеспечением Axiovision 3.1 (Carl Zeiss, Германия). Статистический анализ данных Данные представлены в формате среднее значение ± стандартное отклонение (в ряде случаев стандартная ошибка среднего значения). Оценку Гены & Клетки Том XIII, № 1, 2018 78 ОРИГИНАЛЬНЫЕ ИССЛЕДОВАНИЯ статистической значимости различий проводили с использованием U-критерия Манна-Уитни и программного пакета Statistica 8.0 (Statsoft, сшА). Результаты C-kit+-ПКС были получены из миокарда крыс с помощью метода эксплантной культуры и иммуномагнитной селекции с антителами к маркеру стволовых/прогени-торных клеток c-kit. При анализе иммунофенотипа полученных клеток было обнаружено, что маркер c-kit присутствует на поверхности более чем 90 % клеток (рис. 1А). Маркеры CD34 и CD45 выявлены не были, что исключает костномозговое происхождение популяции этих клеток. Отсутствие маркеров PECAM (CD31), CD34 в выделенной популяции исключает наличие в ее составе и эндотелиальных прогениторных клеток, и клеток зрелого эндотелия [15, 16]. Часть клеток экспрессировала маркеры «стволовости» Oct4, Nanog, что является подтверждением их способности к самообновлению с сохранением их низкодифференцированного состояния в выбранных условиях культивирования (рис. 1 в). С помощью ПЦр в реальном времени было показано, что резидентные ПКС экспрессируют основные проангиогенные факторы роста (VEGF, HGF, ангиопоэтин-1 и ангиопоэтин-2), что может указывать на их ангиостимулирующую активность (рис. 1Б). А Б При изучении мультипотентной дифференцировки С-№-ПКС с помощью культивирования в дифференци-ровочных средах с соответствующими индукторами было обнаружено, что полученные ПКС обладают способностью к мультипотентной дифференцировке (формированию предшественников эндотелильных, гладкомышечных клеток и кардиомиоцитов) (рис. 1 г), что соответствует ранее заявленным свойствам этих клеток [8]. Процесс раннего ремоделирования левого желудочка начинается в первые сут. после возникновения инфаркта миокарда и приводит к повреждению и изменению структурной организации как первичной зоны повреждения, так и зоны сохраненного миокарда лЖ. для воздействия на процессы раннего постинфарктного ремоделирования С-№-ПКС вводили интрамиокардиально в периинфаркт-ную область в острую фазу инфаркта миокарда. Было выявлено, что трансплантированные клетки локализуются в виде обширных скоплений и практически не мигрируют из зоны первичного введения. Только около одной пятой клеток в зоне трансплантации сохраняли способность к пролиферации, о чем свидетельствует экспрессия маркера Ki67 (2061±429 Ю67+-ПКС на 10х103 трансплантированных клеток) (рис. 2а). Также было показано, что часть трансплантированных клеток экспрессировали маркер Pecam (CD31) (рис. 2Б), что косвенно указывает на их дифференцировку в эндотелиальном направлении. CD31 . . 1 ^ ПКС ; ■ ■ ’ Ï ’ . ‘ ' - ; В VEGF HGF Ang 1 Ang 2 C-kit Oct 3/4 Nanog Г CD31 MHC . Phall Gata 6 SMA j Рис. 1. Культуры резидентных ПКС: A - экспрессия c-kit, CD31 и гематопоэтических маркеров (CD34, CD45) проточная цитофлуориметрия; Б - экспрессия генов проангиогенных факторов роста (VEGF, HGF, ангиопоэтин-1 и ангиопоэтин-2); в - иммунофлуоресцентная реакция с антителами к c-kit и маркерам плюрипотентности (Oct %, Nanog); г - иммунофлюоресцентная реакция с антителами к маркерам эндотелия (CD31 - Pecam), кардиомиоцитов (тяжелые цепи миозина) и гладкомышечных клеток (Gata 6, a-SMA) после культивирования в дифференцировочных средах Рис. 2. Характеристика состояния ПКС после трансплантации в миокард, иммунофлуоресцентные реакции с антителами к: А - Ki67; Б - CD31. Трансплантированные клетки мечены флуоресцентным красителем CMDil (красное окрашивание) При количественном анализе площади рубцовой ткани было обнаружено, что достоверных различий между опытной (трансплантация ПКС) и контрольной группами нет (рис. 3). Отсутствие различий в объеме повреждения (зоне рубца) подтолкнули к исследованию второго важного показателя ремоделирования - степени дилатации полости ЯЖ. расширение полости ЯЖ в остром периоде им относится к компенсаторному механизму, поддерживающему систолический объем ЯЖ. При этом оно является специфическим признаком наступления внезапной смерти и развития сердечной недостаточности [17, 18]. Трансплантация ПКС способствовала статистически значимому снижению расширения полости ЯЖ (рис. 3). в основе снижения уровня дилатации ЯЖ могло лежать уменьшение величины трансмурального поражения стенки ЯЖ [19]. Париетальное истончение области инфаркта ЯЖ является следствием снижения сопротивления пораженного миокарда во время систолического напряжения при каждом сокращении и осуществляется за счет нескольких механизмов, включающих растяжение миоцитов с уменьшением их диаметра и скольжение миоцитов по отношению друг к другу. Истончение стенки ЯЖ определяется обширностью поражения, т. е. развитием трансмурального поражения при котором происходит поражение Гены & Клетки Том XIII, № 1, 2018 ОРИГИНАЛЬНЫЕ ИССЛЕДОВАНИЯ 79 клеток во всей толще сердечной стенки. Было обнаружено, что трансплантация ПКС способствует уменьшению распространенности трансмурального повреждения, что является важнейшим показателем восстановления сократительной способности миокарда, уменьшения негативного ремоделирования и снижения риска развития сердечной недостаточности. Размер инфаркта Трансмуральное поражение ЛЖ Индекс дилатации Ш fflf Ш □ ПКС ^Контроль оПКС ^Контроль оПКС «Контроль Рис. 3. Ремоделирования миокарда левого желудочка через 14 сут. после инфаркта миокарда Возможным механизмом снижения уровня дилата-ции ЛЖ и трансмурального поражения является раннее восстановление кровоснабжения в зоне повреждения. Через 14 сут. после трансплантации ПКС количество артериол на мм2 площади в периинфарктной области в опытной группе было значительно выше, чем в группе контроля (рис. 4). Однако эти клетки не несли флуоресцентного маркера (CM Dil) трансплантированных клеток, что исключает возможность их образования из инъецированных клеток. В связи с этим внимание было обращено на клетки эпикарда, которые являются источником васкулогенных клеток-предшественни-ков в эмбриогенезе и постнатальном сердце [20, 21]. Как известно, экспрессия транскрипционного фактора Wt1 - это признак активации эпикарда [22]. A Инъекции ПКС □Контроль вПКС □ Контроль иПКС Рис. 4. Васкуляризация периинфарктной области сердца после трансплантации ПКС и в группе контроля: А - иммунофлуоресцентная реакция миокарда крысы антителами к CD31 (PECAM) (красный) и a-SMA (зеленый), ядра окрашены DAPI; Б - количественная оценка капилляров и артериол в периинфарктной области Было выявлено, что трансплантация ПКС способ-ствовует активации клеток эпикарда и увеличению количества Wt1+-клеток в зоне повреждения (рис. 5А, Б). ПКЭ, экспрессирующие Wt1, появляются в значительном количестве в виде «веретенообразного» слоя на наружной поверхности миокарда (рис. 5А). При этом часть из них мигрирует в субэпикардиальное пространство и миокард. Оценка колокализации маркеров эндотелиальных и гладкомышечных клеток показала, что часть WT1 входят в состав новообразованных сосудов (рис. 5В, Г). При этом совпадения окрашивания с глад-комыщечными клетками наблюдались только в области субэпикарда, c эндотелиальными клетками (маркер PECAM) - как в субэпикарде, так и в миокарде. ' - - ** - Рис. 5. Прогениторные клетки эпикарда через 14 сут. после трансплантации ПКС и в группе контроля: А - иммунофлуоресцентная реакция ткани миокарда с антителами к Wt1 (контрольная группа (слева), трансплантация ПКС (справа); Б - количественная оценка Wt1 +-клеток в группе контроля и после трансплантации ПКС; В - иммунофлуоресцентная реакция ткани миокарда с антителами к Wt1 и CD31; Г - иммунофлуоресцентная реакция ткани миокарда с антителами к Wt1 и a-SMA Обсуждение Результаты проведенных исследований позволяют утверждать, что трансплантация c-k^-ПКС оказывает значительное влияние на регенеративные процессы в постинфарктном сердце. Интрамиокардиальное Г SMA Wt1 Гены & Клетки Том XIII, № 1, 2018 80 ОРИГИНАЛьНыЕ ИссЛЕДОВАНИЯ введение клеток способствовало достоверному уменьшению постинфарктного ремоделирования ЛЖ (снижение индекса дилатации, распространенности трансмурального поражения), что может служить важным индикатором снижения риска развития сердечной недостаточности и внезапной сердечной смерти. В основе уменьшения негативного ремоделирования сердца может лежать активация артериогенеза в зоне повреждения [23], индуцированная трансплантацией прогениторных клеток. Основу постнатального васкулогенеза составляют три основных механизма: ангиогенез, связанный с прорастанием зрелых эндотелиальных клеток из существующих сосудов под действием градиента проангиогенных факторов роста [23], неоваскулоге-нез на основе циркулирующих эндотелиальных кле-ток-предшественников (ЭПК) из костного мозга [15] и адаптивный артериогенез, связанный с образованием коллатералей из предсуществующих артериальных анастомозов [23]. Последние исследования показали [24], что после повреждения формирование новых коронарных сосудов происходит исключительно за счет пролиферации предсуществующих эндотелиальных клеток, а не за счет циркулирующих предшественников. Эти результаты указывают на то, что в миокарде содержится эндогенный пул клеток для формирования новых сосудов. Источником этих клеток могут быть резидентные ПКс, которые локализуются в специализированных «васкулярных клеточных нишах» [25] и после повреждения способны дифференцироваться в направлении эндотелиальных, гладкомышечных клеток и перицитов [26]. В нашем исследовании после трансплантации ПКс сохранялись в миокарде, однако формировали лишь небольшое количество сосудов в зоне повреждения. Причиной ограниченной васкулогенной дифференцировки ПКс может служить их повреждение и массивная гибель при трансплантации в зону ишемии [27]. следовательно, должен существовать другой источник эндотелиальных клеток для формирования значительного количества сосудов в зоне повреждения de novo. Вероятным источником эндотелиальных клеток является эпикард. Эпикард представляет собой тонкое пленчатое образование, покрывающее миокард предсердий и желудочков и сформированное из крупных полигональных клеток плоского эпителия (мезо-телия) и сети коллагеново-эластических волокон. В период эмбрионального развития прогениторные клетки эпикарда (ПКЭ) взаимодействуют между собой, формируют сосудистое сплетение, которое впоследствии разрастается, соединяется с аортой, образуя коронарную сеть будущего сердца [28]. ПКЭ, характеризующиеся экспрессией маркера Wt1, развиваются из мезотелия эпикарда в результате эпителиально-мезенхимального перехода, обусловленного сложными биохимическими перестройками клеток эпикарда, приводящими к приобретению ими мезенхимального фенотипа, устойчивости к апоптозу, повышению миграционных, адгезивных свойств и секреторной активности. Недавно было показано, что этот процесс происходит не только в эмбриональном, но и в постнатальном сердце млекопитающих. Ученые доказали, что эпителиально-мезенхимальный ЛИТЕРАТУРА 1. Cochain C., Channon K.M., Silvestre J.S. Angiogenesis in the infarcted myocardium. Antioxid. Redox Signal. 2013; 18(9): 1100-13. 2. Shah A.M., Mann D.L. In search of new therapeutic targets and strategies for heart failure: recent advances in basic science. Lancet 2011; 378(9792): 704-12. 3. Battegay E.J. Angiogenesis: mechanistic insights, neovascular diseases, and therapeutic prospects. J. Mol. Med. 1995; 73(7): 333-46. переход вызывает образование ПКЭ и их дифференци-ровку в эндотелиальные клетки, формирующие в миокарде новые сосуды [29]. При этом клетки эндотелия секретируют ростовые факторы, запускающие диф-ференцировку ПКЭ в перициты и гладкомышечные клетки, инициирующие сборку и стабилизацию новых сосудов [30]. Мы обнаружили, что трансплантация ПКс способствует активации пула WtV-ПКЭ, формированию сосудистых предшественников и стимуляции артериогенеза в зоне повреждения. Вероятным механизмом активации эпикарда и стимуляции ангиогенеза является секреция трансплантированными клетками проангиогенных факторов роста, а также экзосом/ микровезикул. В нашем исследовании было показано, что ПКс крысы экспрессируют гены проангиогенных факторов роста: HGF, VEGF, ангиопоэтин-1 и ангио-поэтин-2 (рис. 1Б). Кроме того, они продуцируют значительное количество различных цитокинов, включая хемокины (TCA-3, SDF-1 ), сосудистые факторы роста (VEGF, эритропоэтин, bFGF, остеопонтин, SCF) и факторы-регуляторы дифференцировки сердца (Activin A, Dkk homolog-1, TGF-ß), секретируемых в виде белков или упакованных в микровезикулы, свойства которых потенциально могут объяснить терапевтические эффекты ПКс [31]. Известно, что паракринные факторы ПКс, такие как Ang-1, bFGF, HGF, IGF-1, PDGF, SCF, SDF-1 и VEGF, обладают кардиозащитными свойствами и способны стимулировать неоваскуляризацию миокарда [32]. Факторы, продуцируемые трансплантированными ПКс, также могут быть сигналами-индукторами эпителиально-мезенхимального перехода клеток эпикарда. Это прежде всего факторы роста сосудистого эндотелия (VEGF) [33], тромбоцитарный фактор роста PDGF [34, 35], трансформирующий фактор роста бета (TGF beta) [36], морфогенетические белки костной ткани (BMP) [37]. Эти факторы, упакованные в везикулы не только активируют эпителиально-мезенхимальный переход, но и способствуют миграции эндотелиальных клеток-предшественников и подавлению апоптоза [38]. Таким образом, c-k^-ПКс выступают в качестве важного регулятора процессов васкуляризации постин-фарктного сердца. Наряду со способностью ПКс дифференцироваться в эндотелиальном направлении они, вероятно, за счет обмена экзосомами/микровезикулами и действия проангиогенных ростовых фактором, могут активировать пул прогениторных клеток эпикарда и стимулировать ангиогенное поведение эндотелиальных клеток. Это ведет к формированию новых сосудов в зоне повреждения. Раннее формирование сосудов в зоне рубца улучшает трофику ишемизированной ткани, обеспечивая выживаемость кардиомиоцитов и клеток сосудов, уменьшение негативного ремоделирования и прогрессирования сердечной недостаточности.作者简介
K. Dergilev
Russian Cardiology Research and Production Complex
Email: doctorkote@gmail.com
Z. Tsokolaeva
Russian Cardiology Research and Production Complex
I. Beloglazova
Russian Cardiology Research and Production Complex; M.V. Lomonosov Moscow State University
E. Zubkova
Russian Cardiology Research and Production Complex; M.V. Lomonosov Moscow State University
M. Boldyreva
Russian Cardiology Research and Production Complex; M.V. Lomonosov Moscow State University
E. Ratner
Russian Cardiology Research and Production Complex
D. Diykanov
M.V. Lomonosov Moscow State University
M. Menshikov
Russian Cardiology Research and Production Complex
E. Parfenova
Russian Cardiology Research and Production Complex; M.V. Lomonosov Moscow State University
参考
- Cochain C., Channon K.M., Silvestre J.S. Angiogenesis in the infarcted myocardium. Antioxid. Redox Signal. 2013; 18(9): 1100-13.
- Shah A.M., Mann D.L. In search of new therapeutic targets and strategies for heart failure: recent advances in basic science. Lancet 2011; 378(9792): 704-12.
- Battegay E.J. Angiogenesis: mechanistic insights, neovascular diseases, and therapeutic prospects. J. Mol. Med. 1995; 73(7): 333-46.
- Meoli D.F., Sadeghi M.M., Krassilnikova S. et al. Noninvasive imaging of myocardial angiogenesis following experimental myocardial infarction. J. Clin. Invest. 2004; 113(12): 1684-91.
- Uchida Y., Yanagisawa-Miwa A., Nakamura F. et al. Angiogenic therapy of acute myocardial infarction by intrapericardial injection of basic fibroblast growth factor and heparin sulfate: an experimental study. Am. Heart J. 1995; 130(6): 1182-8.
- Gyöngyösi M., Wojakowski W., Lemarchand P. et al. Meta-analysis of cell-based CaRdiac stUdiEs (ACCRUE) in patients with acute myocardial infarction based on individual patient data. Circ. Res. 2015; 116(8): 1346-60.
- Scimia M.C., Gumpert A.M., Koch W.J. Cardiovascular gene therapy for myocardial infarction. Expert Opin. Biol. Ther. 2014; 14(2): 183-95.
- Bearzi C., Rota M., Hosoda T. et al. Human cardiac stem cells. PNAS USA 2007; 104(35): 14068-73.
- Bolli R., Chugh A.R., D'Amario D. et al. Cardiac stem cells in patients with ischaemic cardiomyopathy (SCIPIO): initial results of a randomised phase 1 trial. Lancet 2011; 378(9806): 1847-57.
- Fazel S., Cimini M., Chen L. et al. Cardioprotective c-kit+ cells are from the bone marrow and regulate the myocardial balance of angiogenic cytokines. J. Clin. Invest. 2006; 116(7): 1865-77.
- Kajstura J., Urbanek K., Perl S. et al. Cardiomyogenesis in the adult human heart. Circ. Res. 2010; 107(2): 305-15.
- Dergilev K.V., Makarevich P.I., Tsokolaeva Z.I. et al. Comparison of cardiac stem cell sheets detached by Versene solution and from thermo-responsive dishes reveals similar properties of constructs. Tissue Cell 2017; 49(1): 64-71.
- Traktuev D.O., Tsokolaeva Z.I., Shevelev A.A. et al. Urokinase gene transfer augments angiogenesis in ischemic skeletal and myocardial muscle. Mol. Ther. 2007; 15(11): 1939-46.
- Hochman J.S., Choo H. Limitation of myocardial infarct expansion by reperfusion independent of myocardial salvage. Circulation 1987; 75(1): 299-306.
- Asahara T., Murohara T., Sullivan A. et al. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997; 275(5302): 964-7.
- Lin C.S., Lue T.F. Defining vascular stem cells. Stem Cells Dev. 2013; 22(7): 1018-26.
- Meizlish J.L., Berger H.J., Plankey M. et al.Functional left ventricular aneurysm formation after acute anterior transmural myocardial infarction. Incidence, natural history, and prognostic implications. N. Engl. J. Med. 1984; 311(16): 1001-6.
- Xiao Y., Ding L., Chen H. et al. Grain-Moxibustion may Protect Myocardium by Reducing Oxidative Stress in Doxorubicin-induced Cardiomyopathy Rats. Zhen Ci Yan Jiu. 2016; 41(6): 502-8.
- Paul A., Mitra A., Kohli V. et al. Anaesthetic challenges for device closure of post-infarct ventricular septal defect with coronary angioplasty. Ann. Card. Anaesth. 2003; 6(1): 52-5.
- Duim S.N., Smits A.M., Kruithof B.P. et al. The roadmap of WT1 protein expression in the human fetal heart. J. Mol. Cell. Cardiol. 2016; 90: 139-45.
- Duim S.N., Kurakula K., Goumans M.J. et al. Cardiac endothelial cells express Wilms' tumor-1: Wt1 expression in the developing, adult and infarcted heart. J. Mol. Cell. Cardiol. 2015; 81: 127-35.
- Xiang F.L., Liu Y., Lu X. et al. Cardiac-specific overexpression of human stem cell factor promotes epicardial activation and arteriogenesis after myocardial infarction. Circ. Heart Fail. 2014; 7(5): 831-42.
- Carmeliet P., Jain R.K. Angiogenesis in health and disease. Nat. Med. 2003; 9: 653-60.
- He L., Huang X., Kanisicak O. et al. Preexisting endothelial cells mediate cardiac neovascularization after injury. J. Clin. Invest. 2017; 127(8): 2968-81.
- Gômez-Gaviro M.V., Lovell-Badge R., Fernandez-Avilés F. et al. The vascular stem cell niche. J. Cardiovasc. Transl. Res. 2012; 5(5): 618-30.
- Bearzi C., Leri A., Lo Monaco F. et al. Identifikation of a coronary vascular progenitor cell in the human heart. PNAS USA 2009; 106(37): 15885-90.
- Hamdi H., Furuta A., Bellamy V. et al. Cell delivery: intramyocardial injections or epicardial deposition? A head-to-head comparison. Ann. Thorac. Surg. 2009; 87(4): 1196-203.
- Bogers A.J., Gittenberger-de Groot A.C., Poelmann R.E. et al. Development of the origin of the coronary arteries, a matter of ingrowth or outgrowth? Anat. Embryol. (Berl.) 1989; 180(5): 437-41.
- Reese D.E., Mikawa T., Bader D.M. Development of the coronary vessel system. Circ. Res. 2002; 91(9): 761-8.
- Van den Akker N.M., Winkel L.C., Nisancioglu M.H. et al. PDGF-B signaling is important for murine cardiac development: its role in developing atrioventricular valves, coronaries, and cardiac innervation. Dev. Dyn. 2008; 237(2): 494-503.
- Tang Y., Wu X., Lei W. et al. TGF-beta1-induced migration of bone mesenchymal stem cells couples bone resorption with formation. Nat. Med. 2009; 15(7): 757-65.
- Chimenti I., Smith R.R., Li T.S. et al. Relative roles of direct regeneration versus paracrine effects of human cardiosphere-derived cells transplanted into infarcted mice. Circ. Res. 2010; 106: 971-80.
- Tomanek R.J., Ratajska A., Kitten G.T. et al. Vascular endothelial growth factor expression coincides with coronary vasculogenesis and angiogenesis. Dev. Dyn. 1999; 215(1): 54-61.
- Folkman J., D'Amore P.A. Blood vessel formation: what is its molecular basis? Cell 1996; 87(7): 1153-5.
- Kaminski W.E., Lindahl P., Lin N.L. et al. Basis of hematopoietic defects in platelet-derived growth factor (PDGF)-B and PDGF beta-receptor null mice. Blood 2001; 97(7): 1990-8.
- Austin A.F., Compton L.A., Love J.D. et al. Primary and immortalized mouse epicardial cells undergo differentiation in response to TGFbeta. Dev. Dyn. 2008; 237(2): 366-76.
- Eisenberg L.M., Markwald R.R. Molecular regulation of atrioventricular valvuloseptal morphogenesis. Circ. Res. 1995; 77(1): 1-6.
- Vrijsen K.R., Sluijter J.P., Schuchardt M.W. et al. Cardiomyocyte progenitor cell-derived exosomes stimulate migration of endothelial cells. J. Cell. Mol. Med. 2010; 14: 1064-70.
补充文件
