







Use of morphoj soft package for phenotypic and genotypic variety testing (English oak case study)

- Authors: Baranov S.G1
-
Affiliations:
- Vladimir State University (VlSU)
- Issue: Vol 14, No 4 (2016)
- Pages: 3-13
- Section: Articles
- URL: https://journals.eco-vector.com/ecolgenet/article/view/5984
- DOI: https://doi.org/10.17816/ecogen1443-13
- ID: 5984

Cite item
Full Text

Abstract
Summary: Background. Some special characters in asymmetry of shape were studied in leaf plate English oak. Materials and methods. In 6 random populations generalized Procrustes analysis showed the mix of fluctuating asymmetry (FA) and directional asymmetry (DA). Results. One population showed pure FA. Covariance symmetric matrix reveals more variance in comparison to matrix asymmetry showed variation bilaterally symmetrical landmarks. Correlation analysis of covariance symmetric matrix and matrix asymmetry containing the coordinates of landmarks after the permutation showed a weak positive correlation coefficient (0.25, p < 0.01). Conclusion. The coefficients of variation of the factors “side”and “side × population” among 6 populations were respectively 48.24% and 44.93% that evidenced on equal variance of both types of asymmetry or slightly increased directional asymmetry. It is noted that the cluster analysis the matrix asymmetry can be used to search genotypic variability.
Full Text
Флуктуирующая асимметрия (ФА) — это один из видов билатеральной асимметрии. Она характеризуется незначительным и статистически незначимым отклонением от нуля разности величин правой и левой частей гомологичного билатерально симметричного признака при нормальном распределении этой разности [23, 28]. По современным представлениям, ФА относится к определенному виду изменчивости — реализационной, случайной или флуктуационной [8, 9, 25].
В экологическом мониторинге для определения величины ФА, как правило, используют гомологичные билатерально симметричные друг другу структуры. У насекомых это могут быть ворсинки на теле, жилки и ячеистые структуры на крыльях. У представителей ихтиофауны измеряют число лучей в брюшных плавниках, а также число чешуй в боковой линии. У земноводных используют число полос и пятен на левых и правых конечностях, у птиц — длину рулевых перьев.
В антропометрии принимают во внимание некоторые мерные характеристики, например длину костей конечностей. Обилие признаков вызывает методические затруднения в тестировании величины ФА.
Статистически значимые результаты величины флуктуирующей асимметрии относят к характеристикам популяционной изменчивости.
Дискуссионным остается вопрос о том, что показывает уровень ФА. Предполагают, что отклонения от строгой асимметрии указывают на случайные нарушения развития. Механизмы таких нарушений остаются малоизученными.
Ряд проведенных исследований позволил предложить шкалу стабильности развития с учетом величины индекса ФА или близкого к нему коэффициента стабильности развития [5, 6]. Вместе с тем продолжается совершенствование методов по оценке величины уровня ФА, так же как и поиск биоиндикаторных видов, чувствительных к изменению ФА. Большинство авторов полагает, что отклонение в стабильности развития в популяциях вызывает повышение ФА [5, 15]. Спорным является вопрос о взаимодействии различных по происхождению факторов. В литературных источниках скорее отсутствуют сведения об адаптивном характере повышенной флуктуирующей асимметрии. В то же время отсутствуют данные о снижении серьезных функциональных характеристик при повышенной величине ФА. Древесные биоиндикаторные виды представляют интерес для исследователя из-за фиксированного местонахождения и способности к многолетней реакции на стресс, например вызываемой биоаккумуляцией токсинов. Проведено также изучение влияния климатических особенностей и освещенности на морфологические особенности и стабильность развития, в том числе с использованием популяций некоторых видов дуба [16, 21, 29].
Обычно в экологическом мониторинге используют традиционные методы определения ФА на основе сравнения дисперсий отдельно для правых и левых гомологичных структур.
Различные билатерально симметричные признаки обладают неодинаковой изменчивостью в величине ФА. При этом зачастую проявляется асимметрия с четким преобладанием либо правых, либо левых структур, то есть направленная асимметрия (НА). Поскольку именно ФА считается показателем нестабильности развития, признаки с четкой НА не используются в интегральном экологическом мониторинге [23]. Вместе с тем присутствие НА в смеси с ФА и способность направленной асимметрии к наследованию вызывают интерес именно к этому виду асимметрии [12, 15, 27].
Альтернативным методом принято считать метод геометрической морфометрии [7, 17, 18, 20, 26]. При этом принимаются во внимание метки, расставляемые на билатерально симметричных структурах. Судят об отклонении этих меток от консенсусных точек центроидной фигуры, которая строится в результате усреднения положения меток в системе декартовых координат, и оценивается величина ФА формы того или иного органа или целого организма. Величину ФА определяют в двухфакторном смешанном дисперсионном анализе по величине среднего квадрата дисперсионных остатков взаимодействия двух факторов: «образец» (рандомный) и «сторона» (фиксированный). Первый фактор обозначается кодовыми значениями, соответствующими уровню изменчивости популяции, особи или органа. Фактор «сторона» обозначается только двумя кодовыми значениями («правое» и «левое»). Рассчитывают величины дисперсионных остатков в дисперсионной модели, то есть величины отклонения от консенсусной симметричной фигуры. Величина среднего квадрата фактора «сторона» указывает на присутствие генотипического эффекта направленной асимметрии. По данным многих источников, направленная асимметрия детерминируется несколькими генами по принципу эпистатического эффекта и проявляется примерно у 10 % некоторых мерных признаков, в том числе у листовых пластин древесных растений, например у липы и дуба [10, 11, 15, 17, 18].
В настоящее время популяционная экология активно развивается в области феногенетического мониторинга с использованием фенотипических признаков, служащих феногенетическими маркерами [2, 3]. Известно, что в дисперсионном анализе направленная асимметрия в смеси с ФА дает нежелательный «скос» (bias) в величине ФА. Это обстоятельство не мешает определять величину НА, однако фенотипический эффект флуктуирующей асимметрии остается недоступным для определения [12, 27].
Листовые пластины дуба черешчатого (Quercus robur) имеют особенность в расположении билатерально симметричных элементов. Лопасти в листовой пластине дуба черешчатого не всегда являются парными, то есть на противоположных сторонах может находиться неодинаковое число лопастей. Это объясняется тем, что в процессе формирования пластин на растение действует комплекс факторов. Один из них — освещенность — играет роль в формировании микросреды в различных ярусах фитоценоза в течение онтогенеза дерева. Немаловажное значение имеет пространственная физико-химическая неоднородность совокупности биоценоза.
По сравнению с другими древесными видами у дуба может быть от трех и более выраженных лопастей с неодинаковым числом их на обеих сторонах пластины. Для тестирования асимметрии общепринятым считается подбор парных признаков, не обладающих выраженной асимметричностью, во избежание отклонения разности правое — левое от нормального распределения.
В работах по определению ФА листовых пластин дуба черешчатого рекомендуются различные признаки от одного до девяти, включая такие, как длина черешка, число основных лопастей, лопастей 2-го порядка, расстояние от синусов до средней жилки. Зачастую некоторые признаки могут отсутствовать, а оставшиеся обладать непараметрическим распределением величин разностей, а также обладать направленной асимметрией или антисимметрией. Последний вид асимметрии проявляется при бимодальном распределении разности «правое минус левое» и характеризуется отрицательным эксцессом выборки.
Таким образом, форма и количественные характеристики признаков листовых пластин дуба черешчатого сильно варьируют в связи с генетически закрепленной нормой реакции, поэтому для экологического мониторинга выбирают пластины с набором выраженных билатерально симметричных структур, которые могут считаться метрическими гомологичными парными признаками при традиционном методе анализа.
В предыдущих работах была получена корреляционная зависимость между величиной индекса ФА, полученного линейными измерениями, и величиной индекса ФА, полученного методом геометрической морфометрии. Такая корреляция не может считаться обязательной, а зависит от величины линейных признаков, вносящих больший вклад в форму билатерально симметричных половин [10]. Величины ФА и стабильность/нестабильность развития зависели от сочетания факторов, среди которых существенными были: автовыбросы, промышленные выбросы и высота рельефа, а климатический фактор был значимым во временной протяженности наблюдения.
Изучение соотношения генотипического и фенотипического эффектов в популяциях дуба проводилось лишь косвенно в зависимости от местообитания или климатических особенностей с использованием традиционных линейных методов определения ФА и НА [13, 14]. Большинство работ по феногенетике на основе метода геометрической морфометрии выполнено при изучении популяций грызунов и насекомых [1–4].
Целью настоящей работы было тестирование уровня фенотипической и генотипической изменчивости формы листовой пластины дуба черешчатого при относительно нормальных фоновых экологических условиях. Для тестирования фенотипической изменчивости использовался его средовый компонент, определяемый по величине флуктуирующей асимметрии формы листовой пластины. Для тестирования генотипической изменчивости использовался его компонент, определяемый по величине направленной асимметрии формы листовой пластины. Рабочая гипотеза заключалась в следующем: асимметрия как элемент формы листовой пластины включает генотипический и средовый компоненты изменчивости, определяемые методом геометрической морфометрии.
Материалы и методы
Популяции дуба черешчатого занимают во Владимирской области около 9 % от общей площади лесных массивов. Были выбраны 6 ценопопуляций с ареалами, различавшимися по физико-химическим свойствам почвы, характеру рельефа местности (от 94 до 175 м) и биотопическому видовому составу. В 1–2-й пойменных террасах в районе городов Владимир и Ковров преобладали урочища полого наклонной поймы с ландышевыми дубравами на слабоаллювиальных глеевых супесчаных почвах. В районе Селиваново были выраженные подзолистые почвы, формирующиеся хвойными лесами, в Гусь-Хрустальном районе преобладали болотно-торфяные почвы (рис. 1).
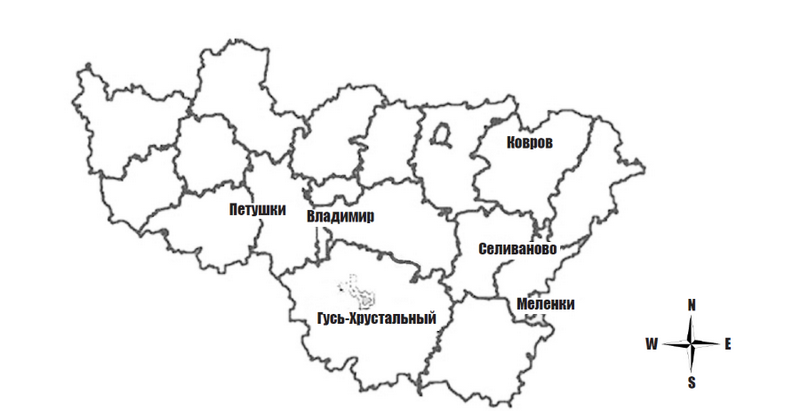
Рис. 1. Расположение популяций (Владимирская обл.)
Fig. 1. Location of populations Quercus robur in Vladimirskay oblast
Сбор листовых пластин дуба черешчатого проводился на протяжении 2014–2015 гг. по общепринятой методике определения стабильности развития В.М. Захарова [5]. Гербаризировались по 100 листовых пластин из каждой популяции от 10 деревьев в условиях относительно одинаковой освещенности с укороченных побегов. Существенным дополнением было то, что а) пластины собирались размером в ширину равным половине листа 3,5–4,5 см и б) отбирались пластины не менее чем с пятью ясно выраженными симметричными лопастями, которые квалифицировались как гомологичные билатерально симметричные признаки.
Листовые пластины были сфотографированы камерой Panasonic DMC-FZ100 и сохранены в формате jpeg. Каждый лист фотографировался отдельно, линии сетки использовались как направляющие для получения по возможности вертикального изображения, для этого рахис (основная вертикальная жилка) при съемке совмещался параллельно с линией сетки. Для нанесения меток (ландмарок) и последующей обработки результатов использовался пакет программ TPS (Rohlf, 2004). Метки расставлялись двукратно для нахождения ошибки измерения и классифицировались как гомологичные метки II типа, так как представляли собой парные точки в местах наибольшей кривизны окончания лопастей или межлопастных синусов (рис. 2).

Рис. 2. a) Десять пар меток для определения флуктуирующей асимметрии, 11–12-я пара меток лежит на оси симметрии; отображение совокупности меток в прокрустовом пространстве ковариационной симметричной матрицы (b) и ковариационной матрицы асимметрии (c). Темным цветом показаны метки, полученные в результате усреднения. Матрица симметрии обладает большей дисперсией из-за неравномерного расположения латеральных лопастей и лопастных пазух. Матрица асимметрии отражает вариацию в расположении меток в левой и правой половинах
Fig. 2. a) 10 pair of landmarks for testing FA; the landmarks represented on a Procrustes fit in symmetric matrix (b) and in matrix of asymmetry (c). Black dots show the landmarks after aliment by superimposition. Symmetric matrix possesses more variance due to variety in lobes and sinuses. Matrix of asymmetry reflects variation of landmarks in left and right sides
Для тестирования обоих видов асимметрии использовался метод прокрустового анализа (Procrustes ANOVA) как аналога двухфакторного дисперсионного смешанного анализа (образец × сторона), который применяется для тестирования флуктуирующей асимметрии мерных и счетных признаков. Прокрустово пространство представляет собой область, ограниченную размером центроида. Прокрустово выравнивание включает операцию зеркального отображения левой и правой сторон, усреднения положения точек и изометрического поворота каждого образца для максимально возможного совмещения образуемых фигур. Подробно о методе геометрической морфометрии изложено в обзорах и руководствах по применению этого метода [7, 20, 26].
В настоящей работе была использована программа MorphoJ1.06d [19], доступная на сайте www.morphometrics.org. Общий файл образца популяции состоял из двухсот файлов (100 листовых пластин × двукратная повторность). Таким образом, строилась таблица 200 × 4, первый столбец содержал координаты меток соответствующих изображений и служил идентификатором (identificator), другие содержали кодовые значения для факторов «популяция», «дерево» и «лист». После создания прокрустова пространства (Procrustes fit) было выполнено тестирование индивидуальной изменчивости на каждом биосистемном уровне. Величина флуктуирующей асимметрии определялась величиной среднего квадрата MS и величиной критерия F Гудолла, оценивающего взаимодействие одного из рандомных факторов — «популяция», «дерево» или «лист» — с фиксированным фактором «сторона».
Были использованы ковариационные матрицы симметрии и антисимметрии, содержащие координаты меток. Канонический вариационный анализ использовался для анализа формы при поиске межгрупповых различий. Анализ главных компонент использовался для анализа различий в форме отдельно взятой группы (популяций, особей деревьев или совокупности листовых пластин).
Для тестирования антисимметрии, третьего вида билатеральной асимметрии, обладающего бимодальным распределением гистограммы частот разности величин правого и левого признаков и отрицательным значением эксцесса, в программе MorphoJ предусмотрено пермутационное размножение выборок, нормализующих их распределение [15, 23]. В предыдущих исследованиях, проведенных традиционным методом, такие свойства в мерных признаках выявлены не были [10]. Вспомогательными программами служили пакеты PAST 3.03 и STATISTICA 10.
Результаты
Ошибка измерения
Листовая пластина представляла единицу измерения и истинную репликацию, так как каждая пластина измерялась дважды. Такая трактовка может представляться критичной [22], учитывая мнение, по которому настоящая, или истинная, репликация измеряется при повторных сборах, то есть листья собираются повторно и выполняются двукратные измерения каждого. Автором принята во внимание точка зрения Клингеберга [10] о двух видах ошибки: оцифровывания и измерения (нанесения меток). При этом проводится повторная съемка с двукратным нанесением меток на каждое изображение. В нашем случае ошибка съемки была менее 1 %, а ошибка измерения от 1 до 10 % от величины среднего квадрата взаимодействия факторов «сторона × образец». Снижение ошибок достигалось с помощью операций масштабирования при работе с экранным дигитайзером TPSdig. Согласно данным Palmer, Strobeck (2003), такой размер ошибки не вызывает искажения величины флуктуирующей асимметрии.
Фенотипический и генотипический компоненты изменчивости совокупности популяций
Сначала был поставлен общий (генерализованный) прокрустов дисперсионный анализ для тестирования НА и ФА для всей совокупности (пула) популяций.
Фактор центроидного размера (корень квадратный из суммы квадратов расстояний от ландмарок до центра изображения) был статистически значимым (F = 260,75; p < 0,0001), что говорило об аллометрическом влиянии величины листовой пластины на их форму. Фактор «популяции» показал высокую статистическую значимость, что свидетельствовало о различии в асимметрии формы листовых пластин в пуле популяций (F = 18,85; p < 0,0001; см табл. 1).
Фактор «сторона», указывающий на направленную асимметрию, также показал статистическую значимость на всех трех биосистемных уровнях (р < 0,0001). Взаимодействие факторов статистически значимо различалось как на популяционном уровне, на уровне дерева как образца генотипа, так и на уровне листовой пластины как фотосинтезирующего органа (во всех случаях p < 0,0001).
Таблица 1. Результаты генерализованного прокрустова анализа Generalized Procrustes ANOVA results
Источник вариации | SS | MS | df | F |
Размер центроида, совокупность популяций | 2,39E + 09 | 4,79E + 08 | 5 | 260,75*** |
Популяции | 2,645 | 0,026 | 100 | 18,85*** |
Сторона | 0,098 | 0,005 | 20 | 3,51*** |
Популяции × сторона | 0,140 | 0,001 | 100 | 1,79*** |
Размер центроида, дерево | 5,36E + 09 | 45449914 | 118 | 422,01*** |
Дерево | 12,969 | 0,005 | 2360 | 5,35*** |
Сторона | 0,098 | 0,005 | 20 | 4,79*** |
Дерево × сторона | 2,426 | 0,001 | 2360 | 1,6*** |
Размер центроида, лист | 5,5E + 09 | 6423502 | 857 | 182,41*** |
Лист | 35,389 | 0,002 | 17140 | 2,44*** |
Сторона | 0,098 | 0,005 | 20 | 5,81*** |
Лист × сторона | 14,510 | 0,001 | 17140 | 4,43*** |
Примечания: SS — сумма квадратов; MS — средний квадрат фактора сторона; df — степень свободы; F – критерий Гудолла; *** p < 0,0001 | ||||
По результатам обобщенного прокрустова анализа для объяснения изменчивости формы были созданы ковариационные матрицы симметрии и асимметрии с координатами меток (см. рис. 2).
При тестировании статистической значимости матрицы симметрии была использована нулевая гипотеза об отсутствии различия в положении структур на каждой половине листовой пластины. Напротив, ковариационная матрица асимметрии отражала изменчивость билатеральных структур с тестированием нулевой гипотезы об отсутствии различия в пространственном положении билатеральных парных меток.
Фактически обе матрицы содержали информацию как о фенотипической, так и о генотипической изменчивости. Исключение представляла матрица асимметрии популяции с «чистой» ФА. В этом случае матрица асимметрии содержала только компонент средового эффекта, исходя из представления об ФА как о фенотипическом проявлении изменчивости [15, 23]. Вместе с тем некоторые работы [24] указывают на возможное присутствие не только общего генотипического, но и генетического, ферментативного и эпигенетического эффекта, влияющего на структуру выборочной частотной характеристики флуктуирующей асимметрии.
Для более полного представления структуры изменчивости формы было проведено парное сравнение двух матриц по 20 главным компонентам с пермутационным размножением выборок и получены коэффициенты парной корреляции для каждой пары главных компонент (табл. 4).
Матрицы различались в общем пуле популяций по главным компонентам (р < 0,0001), а коэффициент корреляции с использованием диагональных блоков показал слабую корреляционную связь между величинами прокрустовых координат (r = 0,25; p < 0,01).
Фенотипический и генотипический компоненты межпопуляционной изменчивости
Для изучения изменчивости формы листовой пластины был выполнен прокрустов дисперсионный анализ для каждой отдельно взятой популяции (табл. 2).
Таблица 2. Результаты прокрустова дисперсионного анализа для популяций Procrustes ANOVA results (for each population)
№ | Популяция | Источниквариации | SS | MS | df | F |
1 | Меленки | Размер центроида | 4461475,28 | 85797,60153 | 52 | 144,79*** |
Лист | 8,496 | 0,003 | 2760 | 2,76*** | ||
Сторона | 0,076 | 0,004 | 20 | 3,43*** | ||
Лист × сторона | 3,076 | 0,001 | 2760 | 6,62*** | ||
2 | Владимир | Размер центроида | 852405,87 | 14696,65288 | 58 | 0,9ns |
Лист | 11,327 | 0,002 | 6260 | 2,51*** | ||
Сторона | 0,027 | 0,001 | 20 | 1,89** | ||
Лист × сторона | 4,507 | 0,001 | 6260 | 2,1*** | ||
3 | Петушки | Размер центроида | 549478,13 | 5494,78134 | 100 | 3,91*** |
Лист | 3,762 | 0,002 | 2380 | 1,91*** | ||
Сторона | 0,039 | 0,002 | 20 | 2,33*** | ||
Лист × сторона | 1,972 | 0,001 | 2380 | 8,72*** | ||
4 | Ковров | Размер центроида | 206188235,40 | 4042906,577 | 51 | 8120,73*** |
Лист | 3,11 | 0,002 | 1420 | 2,8*** | ||
Сторона | 0,05 | 0,003 | 20 | 3,47*** | ||
Лист × сторона | 1,11 | 0,001 | 1420 | 10,18*** | ||
5 | Селиваново | Размер центроида | 206188235,40 | 4042906,577 | 51 | 8120,73*** |
Лист | 1,959 | 0,002 | 1000 | 2,58*** | ||
Сторона | 0,006 | 0,000 | 20 | 0,4ns | ||
Лист × сторона | 0,759 | 0,001 | 1000 | 13,26*** | ||
6 | Гусь-Хрустальный | Размер центроида | 8923323,87 | 171 602,38 | 52 | 256,22*** |
Лист | 4,40 | 0,00 | 3220 | 1,46*** | ||
Сторона | 0,05 | 0,00 | 20 | 2,87*** | ||
Лист × сторона | 3,01 | 0,00 | 3220 | 10,5*** | ||
Примечания: SS — сумма квадратов; MS — средний квадрат; df — степень свободы; F — критерий Гудолла; *** p < 0,0001; ** p < 0,01; ns — p > 0,05 | ||||||
В большинстве популяций величина усредненного центроида оказывала аллометрическое влияние (p < 0,0001) на форму, то есть размер центроида и, следовательно, самой листовой пластины, на индивидуальную изменчивость в асимметрии, что может считаться обычным явлением. Известно, что увеличение размера изучаемого образца прямо влияет на увеличение величины асимметрии, в том числе флуктуирующей [23].
Направленная асимметрия (фактор «сторона»), обладающая компонентом генотипической изменчивости, была статистически значима почти во всех популяциях, кроме популяции № 5, хотя уровень вероятности р был не одинаков. В популяции № 5 (Селиваново) значение F Гудолла дисперсии взаимодействия факторов было наибольшим, соответственно, получена «чистая» флуктуирующей асимметрии на фоне отсутствия НА (p = 0,99).
Был проведен анализ главных компонентов (PCA) и получены прокрустовы координаты первого принципиального компонента, объясняющего более 50 % дисперсии в матрице асимметрии.
Рис. 3 показывает результаты кластерного анализа методом спаренных групп прокрустовых координат первых главных компонент в виде эвклидовых расстояний.
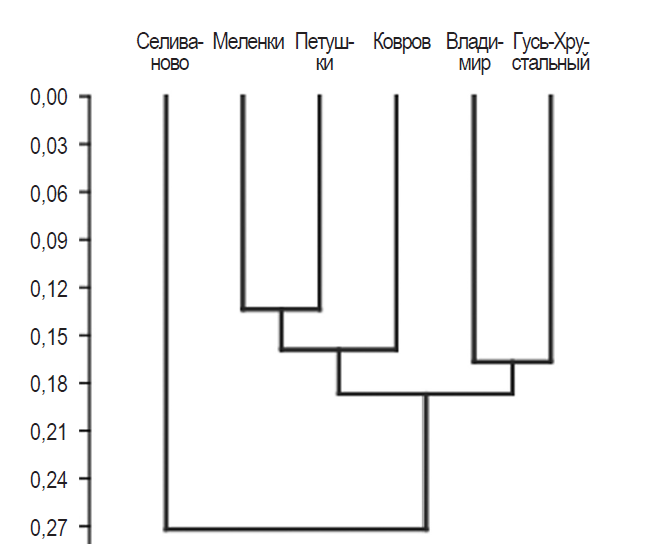
Рис. 3. Объединенная древовидная классификация прокрустовых расстояний шести групп популяций. По оси ординат — эвклидовы расстояния
Fig. 3. Joint classification tree Procrustes distance of six population groups. On the axis of ordinate — Euclidean distance
Эвклидовы дистанции имели статистически значимое различие (табл. 3).
Таблица 3. Значения вероятности р (1000 итераций) для прокрустовых дистанций P-values from permutation tests (10000 permutation rounds) for procrustes distances
Популяция | Гусь-Хрустальный | Меленки | Петушки | Ковров | Владимир | Селиваново |
Гусь-Хрустальный | × | |||||
Меленки | 0,004 | × | ||||
Петушки | 0,002 | 0,006 | × | |||
Ковров | 0,002 | 0,004 | 0,003 | × | ||
Владимир | 0,002 | 0,006 | 0,002 | 0,003 | × | |
Селиваново | 0,001 | 0,005 | 0,002 | 0,002 | 0,003 | × |
Наибольшая статистическая значимость получена при сравнении прокрустовых расстояний популяций Петушки и Меленки (выделено в таблице), что соответствует наибольшему расстоянию между этими местами сбора пластин (более 200 км). Наибольшая величина р (наименьшая статистическая значимость) получена в паре Гусь-Хрустальный — Селиваново, что соответствовало наименьшему географическому расстоянию (около 70 км). Таким образом, смесь ФА + НА показала однородность у близко расположенных популяций, а с удалением популяции различались, что указывает на влияние расстояния на генотипические и фенотипические компоненты изменчивости.
Обсуждение
Листовые пластины дуба черешчатого обладают высоким различием в морфологии с широкой генетически детерминируемой модификационной нормой реакции. Лишь в одной популяции листовые пластины содержали ФА без примеси направленной асимметрии.
В симметричной матрице была получена высокая гетерогенность в значениях главных компонент в сравнении с матрицей асимметрии. Коэффициенты вариации соответственно 10,7 и 9,1 (стандартная ошибка М = ± 0,02; см. также рис. 2, b, c).
Следовательно, латеральные структуры вдоль оси симметрии слева и справа обладали большей изменчивостью, чем билатеральные гомологичные. Была получена зависимость эвклидовых расстояний в кластерном анализе от географической протяженности между местами сборов пластин.
Отмечена корреляция скалярных величин векторов двух видов асимметрии (положительная связь ковариационных матриц). Инициирующим действием предположительно служил эффект генотипической изменчивости формы листовой пластины.
Ковариационные матрицы могут служить скорее визуальным инструментом для оценки миграции меток в контексте общей или фенотипической изменчивости и для поиска коррелятивной зависимости в изменчивости листовой пластины.
Таким образом, геометрическая морфометрия, включавшая канонический вариационный анализ ковариационных матриц и метод главных компонентов, позволяла (хотя и условно) оценивать элементы генотипической и фенотипической изменчивости формы изучаемого объекта.
На взгляд автора, разделение НА и ФА возможно алгебраическим путем при работе со счетными или мерными признаками. Ковариационные матрицы визуально демонстрируют направление векторов, показывающих изменчивость формы, большую долю которой составляет фенотипическая изменчивость, вызванная влиянием среды.
Таблица 4. Коэффициенты парной корреляции асимметричной и симметричной ковариационных матриц Coefficients paired correlation of symmetric matrix and matrix of asymmetry
Главная компонента | Доля объясненной дисперсии, % | Корреляция |
1 | 89,86 | 0,32 |
2 | 7,77 | 0,53 |
3 | 0,77 | 0,37 |
4 | 0,67 | 0,36 |
5 | 0,34 | 0,28 |
6 | 0,27 | 0,33 |
7 | 0,15 | 0,28 |
8 | 0,05 | 0,32 |
9 | 0,04 | 0,20 |
10 | 0,03 | 0,25 |
11 | 0,02 | 0,24 |
12 | 0,01 | 0,17 |
13 | 0,01 | 0,23 |
14 | 0,01 | 0,14 |
15 | 0,00 | 0,13 |
16 | 0,00 | 0,12 |
17 | 0,00 | 0,11 |
18 | 0,00 | 0,08 |
19 | 0,00 | 0,05 |
20 | 0,00 | 0,02 |
Отсутствие статистически значимой направленной асимметрии в популяции Селиваново указывало на отсутствие компонента генотипической изменчивости. Согласно классическому представлению о билатеральной асимметрии [28] структура частотного распределения ФА состоит из средового компонента дисперсии разности величин правых и левых признаков, а диаграмма частот направленной асимметрии состоит из обоих компонентов: средового и генотипического. Тем не менее интерес представляет структура частотной диаграммы выборки с «чистой» ФА.
Распределение главных компонент ковариационной матрицы асимметрии указанных популяций приведено на рис. 4.
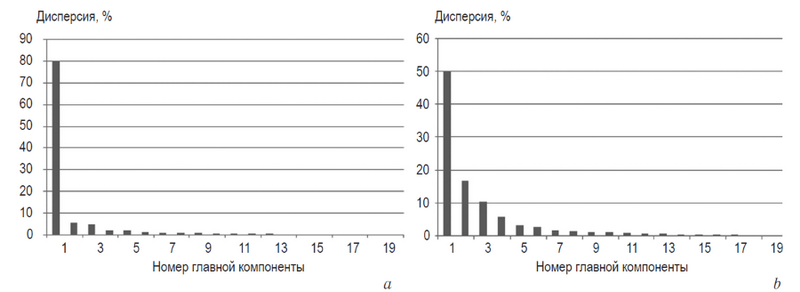
Рис. 4. Диаграмма процентного содержания дисперсии в 20 главных компонентах: a) популяция с чистой флуктуирующей асимметрией, собственное эйген-значение дисперсии 0, 016; b) с примесью направленной асимметрии, собственное эйген-значение дисперсии 0,016. Ось 0x показывает номер главной компоненты
Fig. 4. Diagram of percent values of PC scores (0x axis). a) pure FA (Selivanovo), Eigenvalue variance 0,016; b) FA and DA mix (Vladimir), Eigenvalue variance 0,012. 0x axis shows the number of principal component. 0y axis shows % variance
Как следует из рисунка, выборка с чистой ФА обладала большей величиной собственной дисперсии принципиальных компонентов, чем выборка с отсутствием ФА и присутствием направленной асимметрии. Однако результат по двум выборкам не может считаться показательным. Более того, коэффициенты вариации компонентов «сторона» и взаимодействия «сторона × лист» в изученных популяциях были равны соответственно 48,24 и 44,93 %, что свидетельствовало об одинаковой/почти одинаковой дисперсии двух компонентов дисперсионного анализа. Данные результаты согласуются с результатами, полученными на дрозофилах при изучении соотношения НА и ФА [27].
Величина генотипической составляющей в частотном распределении НА является трудно определяемой величиной. При высоком значении НА можно лишь предположить высокую численность значений генотипической составляющей направленной асимметрии. Механизм действия такого компонента может включать не только эпистатическое действие генов, но и реакции на молекулярно-генетическом субуровне, включая и эпигенетический эффект [24].
Ограничением описанного использования ковариационных матриц и метода геометрической морфометрии в целом является высокая гетерогенность в морфологии листовых пластин. Частотное распределение (и распределение по главным компонентам) флуктуирующей асимметрии может включать как высокую гетерогенность прокрустовых остатков, так и их низкую гетерогенность. Исходя из этого, интерес представляют исследования в сфере изучения гетерогенности структуры ФА в зависимости от стресс-факторов в сравнении с контролем. Кластерный анализ удачно разделил популяции по асимметричности формы (включая смесь ФА и НА), что согласовалось с географическим расположением популяций.
В районе Селиваново преобладают подзолистые почвы супесчаного и песчаного типов, которые формируют определенный гидрологический, минеральный и, соответственно, экологический режим биоценоза, что могло вызвать повышенную величину ФА по сравнению с более плодородными почвами, более типичными и благоприятными для дуба черешчатого.
Тестирование фенотипической и генотипической изменчивости с помощью описанных подходов может применяться при исследовании на различных уровнях биосистем, включая их биомедицинские аспекты.
Благодарности
Автор благодарит сотрудников и студентов ВлГУ за помощь в сборе гербарного материала и статистической обработке первичных данных и заявляет об отсутствии конфликта интересов, вызванных публикацией статьи.
About the authors
Sergey G Baranov
Vladimir State University (VlSU)
Author for correspondence.
Email: bar.serg58@gmail.com
docent, Department of Biological and Geographical Education Russian Federation
References
- Большаков В.Н., Васильев А.Г., Васильева И.А., и др. Сопряженная биотопическая изменчивость ценопопуляций симпатрических видов грызунов на Южном Урале // Экология. – 2015. – № 4. – С. 265–271. [Bol’shakov VN, Vasil’ev AG, Vasil’eva IA, et al. Coupled biotopic variation in populations of sympatric rodent species in the Southern Urals. Russ J of Ecol. 2015;46(4): N.p., Print. (In Russ.)]
- Васильев А.Г., Васильева И.А. Феногенетический мониторинг импактных популяций растений и животных в условиях антропогенного пресса // Научные ведомости БелГУ. – Серия «Естественные науки». – 2009. – Т. 8. URL: http://cyberleninka.ru/article/n/fenogeneticheskiy-monitoring-impaktnyh-populyatsiy-rasteniy-i-zhivotnyh-v-usloviyah-antropogennogo-pressa (дата обращения 24.02.2016). [Vasil’ev AG, Vasil’eva IA. Fenogeneticheskii monitoring impaktnykh populyatsii rastenii i zhivotnykh v usloviyakh antropogennogo pressa. Nauchnye vedomosti BelGU. Seriya: Estestvennye nauki. 2009;8. (In Russ.)]
- Васильев А.Г., Васильева И.А., Большаков В.Н. Эволюционно-экологический анализ закономерностей феногенетической изменчивости гомологичных морфоструктур: от популяций до экологических рядов видов // Экология. – 2010. – Т. 5. – С. 1–8. [Vasilyev AG, Vasilyeva IA, Bol’shakov VN. Evolutionary-Ecological analysis of trends in phenogenetic variation of homologous morphological structures: from populations to ecological series of species. Russ J Ecol. 2010;41(5):365-371. Web. 23 May 2016. (In Russ.)]
- Войта Л.Л., Омелько В.Е., Петрова Е.А. Анализ морфометрической изменчивости и внутривидовой структуры крошечной бурозубки Sorex minutissimus Zimmermann, 1780 (lipotyphla: soricidae) на территории России // Труды Зоологического института РАН. – 2013. – Т. 317. – Вып. 3. – С. 332–351. [Voita LL, Omelko VE, Petroava EA. Analysis of the morphometrics variability and intraspecific structure of Sorex Minutissimus Zimmermann, 1780 (Lipotyphla: Soricidae) in Russia. Acta Theriol. 2013;8:167-179:11-13. (In Russ.)]. doi: 10.4098/at.arch.64-10.
- Захаров В.М., Чубинишвили А.Т. Мониторинг здоровья среды на охраняемых природных территориях. – М.: Центр экологической политики России, 2001. – 148 с. [Zakharov VM, Chubinishvilli AT. Monitoring zdorov’ya sredy na okhranyaemykh prirodnykh territoriyakh. Moscow: Tsentr ekologicheskoi politiki Rossii; 2001. 148 p. (In Russ.)]
- Луговская Л.А., Межова Л.А. Биоиндикация геоэкологических условий с использованием дуба черешчатого (Quercus robur L.) для мониторинга среды // Проблемы региональной экологии. – № 2. – 2012. – С. 65–68. [Lugovskaja LA, Mezhova LA. Bioindikacija geojekologicheskih uslovij s ispol’zovaniem duba chereshchatogo (Quercus robur L.) dlja monitoringa sredy. Problemy regional’noj jekologii. 2012(2):65-68. (In Russ.)]
- Павлинов И.Я., Микешина Н.Г. Принципы и методы геометрической морфометрии // Журнал общей биологии. – 2002. – Т. 63. – № 6. – С. 473–493. [Pavlinov LYa, Mikeshina NG. Principles and methods of geometric morphometrics. Russian Journal of Ecology. 2002;63(6):473-493. (In Russ.)]
- Струнников В.А., Вышинский И.М. Реализационная изменчивость у тyтoвoгo шелкопряда // Проблемы генетики и теории эволюции. – Новосибирск: Наука, 1991. – С. 99–114. [Strunnikov VA, Vyshinskij IM. Realizacionnaja izmenchivost’ u tytovogo shelkoprjada. Problemy genetiki i teorii jevoljucii. Novosibirsk: Nauka; 1991. P. 99-114. (In Russ.)]
- Тиходеев О.Н. Классификация изменчивости по факторам, определяющим фенотип: традиционные взгляды и их современная ревизия // Экол. генетика. – 2013. – Т. 11. – Вып. 3. – С. 79–92. [Tihodeev ON. Klassifikacija izmenchivosti po faktoram, opredeljajushhim fenotip: tradicionnye vzgljady i ih sovremennaja revizija. Jekol. genetika. 2013;11(3):79-92 (In Russ.)]
- Baranov SG, Zykov IE, Fedorova LV. Developmental stability study of Quercus Robur: industrial and abiotic factors influence. Adv Environ Biol. 2014;8(17):102-109.
- Baranov SG. Use of morphogeometric method for study fluctuating asymmetry in leaves Tilia cordata under industrial pollution. Adv Environ Biol. 2014;8(7):2391-2398.
- Costa M, Mateus R, Moura M. Constant fluctuating asymmetry but not directional asymmetry along the geographic distribution of Drosophila antonietae (Diptera, Drosophilidae). Revista Brasileira de Entomologia. 2015;59:337-342. doi.org/10.1016/j.rbe.2015.09.004.
- Fair JM, Breshears DD. Drought stress and fluctuating asymmetry in Quercus undulata leaves: confounding effects of absolute and relative amounts of stress? J Arid Environ. 2005;62(2):235-249.
- Fei Xu, Weihua Guo, Weihong Xu And Renqing Wang. Habitat effects on leaf morphological plasticity in Quercus acutissima. Acta Biologica Cracoviensia Series Botanica. 2008;50(2):19-26.
- Graham John H, Whitesell Mattie J, Fleming Mark II, et al. Fluctuating asymmetry of plant leaves: batch processing with LAMINA and continuous symmetry measures. Symmetry. 2015;7:255-268. doi: 10.3390/sym7010255.
- Hodar Jose A. Leaf fluctuating asymmetry of Holm oak in response to drought under contrasting climatic conditions. J Arid Environ. 2002;52:233-243.
- Klingenberg CP, Duttke S, Whelan S, Kim M. Developmental plasticity, morphological variation and evolvability: a multilevel analysis of morphometric integration in the shape of compound leaves. J Evol Biol. 2012;25:115-129.
- Klingenberg CP. Analyzing fluctuating asymmetry with geometric morphometrics: concepts, methods, and applications. Symmetry. 2015;7843-934. doi: 10.3390/sym7020843.
- Klingenberg CP. MorphoJ: an integrated software package for geometric morphometrics. Mol Ecol Res. 2011;11:353-357. doi: 10.1111/j.1755-0998.2010.02924.x.
- Klingenberg СР, Barluenga M, Meyer A. Shape analysis of symmetric structures: quantifying variation among individuals and asymmetry. Evol. 2002;56:1909-1920. doi: 10.1554/0014-3820.
- Nuche P, Komac B, Camarer JJ, et al. Developmental instability as an index of adaptation to drought stress in a Mediterranean oak. Ecol Indic. 2014;40 May: 68-75.
- Oksanen L. The devil lies in details: reply to Stuart Hurlbert. Oikos. 2004;104:598-605.
- Palmer AR, Strobeck C. Fluctuating asymmetry as a measure of developmental stability: Implications of non-normal distributions and power of statistical tests. Acta Zool Fennica. 191:57-72 Helsinki 30 June 1992.
- Parsons PA. Fluctuating asymmetry: an epigenetic measure of stress. Biol Rev. 1990;65(2):131-145.
- Raj A, Van Oudenaarden A. Nature, nurture, or chance: stochastic gene expression and its consequences. Cell. 2008;135:216-226.
- Rohlf FJ. Shape statistics: procrustes superimpositions and tangent spaces. J Classif. 1999;16:197-223.
- Stige LC, David B, Alibert P. On hidden heterogeneity in directional asymmetry – can systematic bias be avoided? J Evol Biol. 2006;19:492-499. doi: 10.1111/j.1420-9101.2005.01011.x.
- Van Valen L. A study of fluctuating asymmetry. Evol. 1962;16(2):125-142.
- Viscosi V, Cardini A. Leaf morphology, taxonomy and geometric morphometrics: a simplified protocol for beginners. PLoSONE. 2011;6(10):25630. doi: 10.1371/journal.pone.0025630.
Supplementary files



