







Karyotype and inversion polymorphism of natural populations Glyptotendipes glaucus (meigen), 1818 (diptera, chironomidae) from the small reservoirs of Kaliningrad

- Authors: Vinokurova N.V1, Kalinina E.A1, Stol’ E.E1
-
Affiliations:
- Immanuel Kant Baltic Federal University, Kaliningrad
- Issue: Vol 14, No 4 (2016)
- Pages: 41-51
- Section: Articles
- URL: https://journals.eco-vector.com/ecolgenet/article/view/5988
- DOI: https://doi.org/10.17816/ecogen14441-51
- ID: 5988

Cite item
Full Text

Abstract
Full Text
ВВЕДЕНИЕ
Личинки вида Glyptotendipes glaucus Mg., наравне с другими видами сем. Chironomidae, являются неотъемлемыми компонентами водных биоценозов Голарктики, занимая в них различные экологические ниши. G. glaucus распространен в прибрежьях водохранилищ, литорали озер, прудов, в лужах и слабо проточных заросших участках равнинных рек. Личинки способны минировать отмершие макрофиты и древесину, а также являются обрастателями и поселяются на поверхности погруженных предметов (пни, коряги, камни, макрофиты) [1, 2].
Морфологические, экологические и кариологические особенности данного вида являются предметом неизменного научного интереса ученых.
Как известно, популяции G. glaucus характеризуются высоким уровнем хромосомного полиморфизма, который возможно и необходимо учитывать при проведении экологических мониторинговых исследований в условиях возрастающих антропогенных загрязнений [3, 4].
На территории Калининградской области вид G. glaucus малоизучен. Данных по структурно-функциональной организации политенных хромосом и уровне естественного полиморфизма природных популяций G. glaucus в области крайне мало [5], и это исследование дополняет сведения о кариофонде вида. В настоящей работе представлены новые данные по хромосомному полиморфизму популяций G. Glaucus из разных водоемов г. Калининграда, а также произведен сравнительный анализ полученных результатов с более ранними данными из калининградских водоемов и других мест обитания изучаемого вида.
МАТЕРИАЛЫ И МЕТОДЫ
Объектом для исследования особенностей кариотипа G. glaucus послужили 136 личинок IV стадии развития из 5 следующих водоемов г. Калининграда: прудов Чистый и Мельничный, системы прудов Карасевка, озер Пеньковое и Школьное. Количественный сбор проб материала для анализа произведен в июне 2012 г. с корней и погруженных в воду частей прибрежных растений. Личинки были зафиксированы по стандартной методике [6] в смеси 96 % этилового спирта и ледяной уксусной кислоты в соотношении 3 : 1 (фиксатор Карнуа). Фиксатор в емкости с личинками меняли до тех пор, пока он не станет прозрачным.
Временные цитологические препараты политенных хромосом слюнных желез личинок готовили по стандартной ацето-орсеиновой методике [7]. Из 2–3-го грудного сегмента фиксированной личинки в капле 40 % молочной кислоты выделяли слюнные железы и окрашивали орсеином на предметном стекле в течение 30 минут. Затем железы переносили в каплю 50 % молочной кислоты, где их отделяли от секрета железы и разрушали, для того чтобы хромосомы могли быть извлечены и отделены от цитоплазмы. Для получения отчетливого рисунка хромосомных дисков в процессе анализа освобожденные хромосомы накрывали покровным стеклом, а излишки молочной кислоты удаляли фильтровальной бумагой с легким надавливанием. По краю покровного стекла проводили прозрачным лаком, чтобы препятствовать быстрому высыханию препарата. Готовые временные препараты хранили в холодильнике [8].
Картирование хромосом исследуемого вида выполнено по системе Беляниной, Дурновой (1998) [9]. Последовательности дисков в каждом плече обозначали сокращенным символом вида, символом плеча и номером последовательности (к примеру, glaE1).
Фотографии кариотипа G. glaucus были получены с помощью микроскопа Axio Scope.A1 и фотокамеры AxioCamMRc5 (CarlZeiss, Германия).
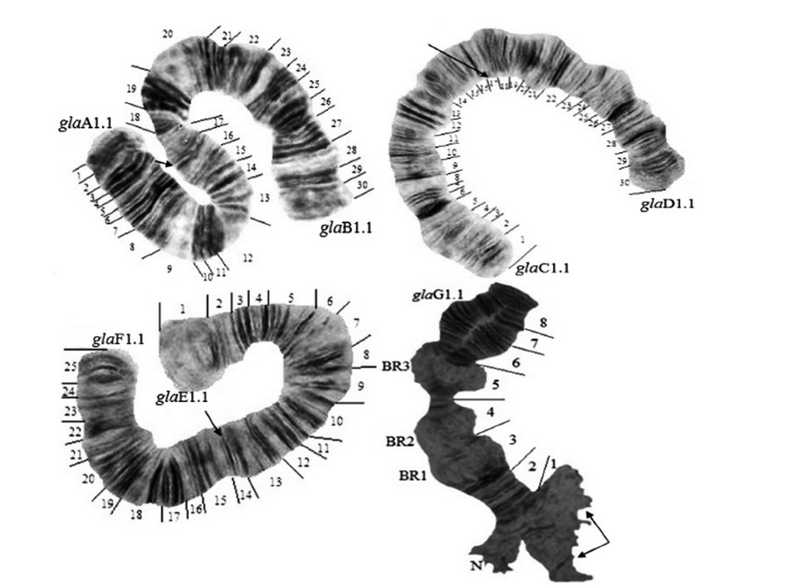
Рис. 1. Политенные хромосомы Glyptotendipes glaucus Mg. из г. Калининграда: glaA1.1, glaB1.1, glaC1.1, glaD1.1, glaE1.1, glaF1.1, glaG1.1 — символы генотипических сочетаний последовательностей дисков в плечах хромосом, BR — кольцо Бальбиани, N — ядрышковый организатор, стрелками указаны центромеры
Fig. 1. Polytene chromosomes of Glyptotendipes glaucus Mg. from Kaliningrad. glaA1.1, glaB2.2, glaC1.1, glaD1.1, glaE1.1, glaF1.1, glaG1.1 – symbols of genotypic combinations of band sequences in chromosomal arms, BR – Balbiani ring, N – nucleolar organizer. Arrows indicate the centromeres
Уровень хромосомного полиморфизма рассчитывался по числу обнаруженных гетерозиготных и гомозиготных инверсий, приходящихся на одну особь. В качестве показателей генотипической структуры исследованных природных популяций личинок были использованы среднее число особей с гетерозиготными инверсиями, количество стандартных и инверсионных последовательностей в популяции, количество геномных комбинаций последовательностей дисков и новый, предложенный нами показатель соотношения числа инверсионных последовательностей к числу геномных комбинаций [10–12]. Для оценки соответствия фактического распределения частот гетерозиготных особей в пределах каждой популяции с теоретически ожидаемым использовали метод χ2 (≤ 0,9) [13].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Структура кариотипа естественных популяций G. glaucus из исследованных водоемов г. Калининграда соответствует ранее описанному кариотипу данного вида [6, 14, 15]. Однако в результате исследования кариотипа калининградских популяций G. glaucus были обнаружены некоторые особенности (рис.1). Так, в плече А имеется одно ядрышко (N1), а в плече Е — два ядрышка (N1 и N2). Хромосома G в районе центромеры имеет одно ядрышко (N) и три кольца Бальбиани (BR1, BR2, BR3) на всем протяжении хромосомы. В плечах С и F также обнаружены пуфы: два ярко выраженных пуфа (P1 и P2) в плече С и один крайне редкий и слабовыраженный пуф (P1) в плече F [16].
При исследовании особенностей структурной организации кариотипа G. glaucus природных популяций водоемов г. Калининграда были установлены частоты стандартных и инвертированных последовательностей дисков (ПД) (табл. 1).
В выборках исследованных популяций было определено 20 инверсионных последовательностей.
Плечо А в исследованных популяциях было представлено тремя последовательностями (glaA1, glaA2, glaA3). Во всех выборках превалировала стандартная последовательность glaA1 (от 0,629 до 1,0) (см. табл. 1). Последовательность glaA2 была обнаружена в гомо- и гетерозиготном состояниях. Последовательность glaA3 была встречена в гетерозиготном состоянии только в системе прудов Карасевка (0,074).
Плечо В. Отмечен высокий уровень полиморфности данного плеча, при анализе которого было выявлено четыре последовательности: glaВ1, glaВ2, glaВ3, glaВ5. Распространенной последовательностью в исследованных популяциях была glaВ1 (от 0,593 до 1,0). Последовательность glaB2 в гомо- и гетерозиготных состояниях зафиксирована в четырех популяциях: в системе прудов Карасевка — 0,296, в пруду Чистом — 0,370, в озере Школьном — 0,365, в пруду Мельничном — 0,407. В пруду Чистом и озере Школьном была обнаружена последовательность glaВ3 в гетерозиготном состоянии, частота которой составила 0,037 и 0,038 соответственно (см. табл. 1). Последовательность glaB5, которая отличается от последовательности glaВ2 инверсией 20 — 19a-d [5], была отмечена в озере Школьном, и частота ее встречаемости составила 0,019 (см. табл. 1).
Плечо С было представлено тремя последовательностями: glaC1, glaC2, glaC3. Стандартная геномная комбинация glaC1.1 преобладала во всех исследованных природных популяциях. В кариотипах личинок из озера Пенькового и пруда Мельничного кроме данной последовательности других не наблюдалось.
Последовательность glaC2 с невысокой частотой была отмечена в двух прудах: Карасевке (0,037) и Школьном (0,019). В популяции из системы прудов Карасевка у одной особи наблюдалась последовательность glaC3 (0,037) (см. табл. 1).
Таблица 1. Частота встречаемости последовательностей дисков в плечах хромосом Glyptotendipes glaucus природных популяций водоемов г. КалининградаFrequencies of band sequences in chromosomal arms of Glyptotendipes glaucus of natural populations from reservoirs of Kaliningrad
Последовательности дисков | Карасевка 13.06.12, n = 27 | Чистый 14.06.12, n = 27 | Школьное 15.06.12, n = 52 | Пеньковое 18.06.12, n = 3 | Мельничный 21.06.12, n = 27 |
glaA1 | 0,629 | 0,926 | 0,885 | + | 0,926 |
glaA2 | 0,296 | 0,074 | 0,115 | – | 0,074 |
glaA3 | 0,074 | – | – | – | – |
Hob | 0,333 | 0,074 | 0,077 | – | 0,074 |
Hex | 0,510 | 0,137 | 0,204 | – | 0,137 |
glaB1 | 0,703 | 0,593 | 0,577 | + | 0,593 |
glaB2 | 0,296 | 0,370 | 0,365 | – | 0,407 |
glaB3 | – | 0,037 | 0,038 | – | – |
glaB5 | – | – | 0,019 | – | – |
Hob | 0,111 | 0,333 | 0,230 | – | 0,296 |
Hex | 0,416 | 0,510 | 0,531 | – | 0,483 |
glaC1 | 0,926 | 1,0 | 0,980 | + | 1,0 |
glaC2 | 0,037 | – | 0,019 | – | – |
glaC3 | 0,037 | – | – | – | – |
Hob | 0,074 | 0 | 0,019 | – | 0 |
Hex | 0,138 | 0 | 0,037 | – | 0 |
glaD1 | 0,963 | 0,926 | 0,827 | + | 1,0 |
glaD2 | – | 0,074 | 0,154 | – | – |
glaD3 | 0,037 | – | 0,019 | – | – |
Hob | 0,037 | 0,074 | 0,154 | – | 0 |
Hex | 0,071 | 0,137 | 0,287 | 0 | |
glaE1 | 1,0 | 1,0 | 0,942 | + | 1,0 |
glaE6 | – | – | 0,058 | – | – |
Hob | 0 | 0 | 0,058 | – | 0 |
Hex | 0 | 0 | 0,109 | 0 | |
glaF1 | 0,963 | 1,0 | 0,942 | + | 1,0 |
glaF2 | – | – | 0,019 | – | – |
glaF3 | 0,037 | – | – | – | – |
glaF7 | – | – | 0,058 | – | – |
Hob | 0,037 | 0 | 0,077 | – | 0 |
Hex | 0,071 | 0 | 0,148 | – | 0 |
glaG1 | 0,629 | 0,852 | 0,750 | + | 0,778 |
glaG4 | 0,370 | 0,148 | 0,250 | – | 0,222 |
Hob | 0,333 | 0,148 | 0,192 | – | 0,222 |
Hex | 0,465 | 0,252 | 0,375 | – | 0,345 |
Примечание: Hob — наблюдаемая гетерозиготность, Hex — ожидаемая гетерозиготность | |||||
Плечо D в основном было представлено стандартной последовательностью glaD1 (от 0,827 до 1,0). В пруду Чистом и озере Школьном была выявлена последовательность дисков glaD2, частота которой составила 0,074 и 0,154 соответственно. По одной особи с последовательностью glaD3 было обнаружено в пруду Карасевка (0,037) и озере Школьном (0,019) (см. табл. 1).
Плечо E имело только стандартный рисунок дисков за исключением особей популяции из озера Школьного, в которой у трех особей наблюдалась последовательность glaE6 + glaF7. Вышеописанная последовательность glaE6 + glaF7 является сочетанием парацентрической glaE6 и перицентрической glaF7 гетерозиготных инверсий в хромосоме IIIEF (табл. 2) [5]. Так как данные инверсии были впервые обнаружены в системе прудов Карасевка и озере Школьном г. Калининграда и встречались только в сочетании, но никогда отдельно, кроме того, затрагивали оба плеча и центромеру, то в дальнейшем ее обозначение предлагается оставить как glaE6 + glaF7.
Плечо F, как и плечо В, личинок исследованных популяций показало более высокий уровень полиморфизма по сравнению с другими плечами и было представлено четырьмя последовательностями (glaF1, glaF2, glaF3, glaF7) с преобладающей во всех выборках стандартной последовательностью glaF1 (от 0,942 до 1,0) (см. табл. 1). Последовательности glaF2 и glaF3 были отмечены у единичных личинок в озере Школьном (0,019) и пруду Карасевка (0,037) соответственно. В природной популяции G. glaucus озера Школьного была идентифицирована последовательность дисков glaF7 (см. табл. 2).
Таблица 2. Локализация инверсионных последовательностей дисков в плечах хромосом Glyptotendipes glaucus Localization of inversion band sequences in chromosomal arms of Glyptotendipes glaucus
Последовательность дисков | Порядок дисков в хромосомах |
glaA1 | 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 |
glaA2 | 1 10 9 8 7 6 5 4 3 2 11 12 13 14 15 16 |
glaA3 | 1 2 3 4 5 6 7 8 9 10 11 13 12 14 15 16 |
glaB1 | 17 18 19 20 21 22 23 24 25 26 27 28 29 30 |
glaB2 | 17 18 20 1921 22 23 24 25 26 27 28 29 20 |
glaВ3 | 17 18 20 192128 27 26 25 24 23 22 29 30 |
glaВ5 | 17 18 20 19n-e 22f-a21a-d19a-d 22g-k 23 24 25 26 27 28 29 30 |
glaC1 | 1 2 3 4 5 6 7 8 9 10 12 13 14 15 16 |
glaC2 | 1 2 3 4 5 6 7 8 9a-c 13e-a 12 11 10 9j-d 13fg 14 15 16 |
glaC3 | 1 11 10 9 8 7 6 5 4 3 2 12 13 14 15 16 |
glaD1 | 17 18 19 20 21 22 23 24 25 26 27 28 29 30 |
glaD2 | 17 18 19 20 21 22 23 29d-a 28 27 26 25 24 29e-g30 |
glaD3 | 17–27 28 29ab 30dcba 29gfedc 30e-I |
glaE1 | 1 2 3 4 5 6 7 8 9 10 11 12 13 14 |
glaE6 + glaF7 | 1 2 3 4 5 6 7 8 9 10 15 i-a K 14 13 12 11 15j-o 16 17 |
glaF1 | 15 16 17 18 19 20 21 22 23 24 25 |
glaF2 | 15a-h 18 17 16 15n-i 19 20 21 22 23 24 25 |
glaF3 | 15a-h 23 22 21 20 19 18 17 16 15n-i 24 25 |
glaF7 | 15 а-i K 14 13 12 11 15 j-o 16 17 18 19 20 21 22 23 24 25 |
glaG1 | 1 2 3 4 5 6 7 8 |
glaG4 | 1 2 3 7 6 5 4 8 |
Примечание: подчеркнуты инвертированные участки | |
Плечо G. В целом во всех популяциях преобладает стандартная последовательность glaG1 (от 0,629 до 1,0) (см. табл. 1). Инвертированная последовательность glaG4 в гетерозиготном состоянии также встречена во всех исследуемых популяциях, кроме озера Пенькового. В двух выборках последовательность glaG4 отмечена в гомозиготном состоянии (система прудов Карасевка и озеро Школьное).
Таким образом, 20 последовательностей дисков хромосом, обнаруженных в исследованных природных популяциях, сформировали 30 геномных комбинаций (табл. 3).
Таблица 3. Геномные комбинации в кариотипах личинок Glyptotendipes glaucus и их абсолютное число в водоемах г. КалининградаGenomic combinations in karyotypes of Glyptotendipes glaucus larvae and their absolute frequency in reservoirs of Kaliningrad
Геномные комбинации | Карасевка | Чистый | Школьное | Пеньковое | Мельничный | Всего |
A11 B11 C11 D11 E11 F11 G11 | 7номные | 13 | 16 | 3 | 15 | 54 |
A11 B25 C11 D11 E11 F11 G11 | – | – | 1 | – | – | 1 |
A11 B23 C11 D11 E11 F11 G11 | – | – | 1 | – | – | 1 |
A11 B22 C11 D11 E11 F11 G11 | 2 | – | 4 | – | – | 6 |
A11 B12 C11 D11 E11 F11 G11 | 2 | 6 | 4 | – | 4 | 16 |
A11 B23 C11 D11 E11 F11 G44 | – | – | 1 | – | – | 1 |
A11 B22 C11 D11 E11 F11 G14 | 2 | 2 | 3 | – | 2 | 9 |
A12 B22 C11 D11 E11 F11 G11 | – | – | 1 | – | 1 | 2 |
A11 B12 C11 D13 E11 F11 G11 | – | – | 1 | – | – | 1 |
A11 B11 C11 D11 E11 F11 G14 | 2 | – | 2 | – | – | 4 |
A22 B11 C11 D11 E11 F11 G11 | – | – | 1 | – | – | 1 |
A11 B22 C12 D11 E11 F12 G11 | – | – | 1 | – | – | 1 |
A12 B11 C11 D11 E11 F11 G11 | 1 | 2 | 2 | – | 1 | 6 |
A11 B11 C11 D11 E16 F17 G14 | – | – | 1 | – | – | 1 |
A11 B11 C11 D12 E11 F11 G14 | – | – | 3 | – | – | 3 |
A12 B11 C11 D11 E11 F11 G14 | 3 | – | 1 | – | – | 4 |
A22 B12 C11 D11 E11 F11 G44 | – | – | 1 | – | – | 1 |
A11 B11 C11 D11 E16 F17 G11 | – | – | 1 | – | – | 1 |
A11 B11 C11 D12 E11 F11 G11 | – | 1 | 2 | – | – | 3 |
A11 B12 C11 D11 E16 F17 G11 | – | – | 1 | – | – | 1 |
A11 B12 C11 D12 E11 F11 G11 | – | 1 | 2 | – | – | 3 |
A22 B11 C11 D13 E11 F11 G14 | 1 | – | – | – | – | 1 |
A13 B11 C12 D11 E11 F11 G11 | 1 | – | – | – | – | 1 |
A12 B12 C11 D11 E11 F11 G14 | 1 | – | – | – | – | 1 |
A12 B11 C11 D11 E11 F11 G44 | 1 | – | – | – | – | 1 |
A11 B22 C11 D11 E11 F13 G11 | 1 | – | – | – | – | 1 |
A13 B11 C11 D11 E11 F11 G11 | 1 | – | – | – | – | 1 |
A12 B11 C13 D11 E11 F11 G11 | 1 | – | – | – | – | 1 |
A11 B12 C11 D11 E11 F11 G14 | – | 1 | – | – | 4 | 5 |
A11 B13 C11 D11 E11 F11 G14 | – | 1 | – | – | – | 1 |
Примечание: полужирным шрифтом выделены инверсии | ||||||
Следует отметить, что в изученных природных популяциях обнаружено небольшое число особей со стандартным кариотипом.
Преобладающая во всех выборках в гомо- и гетерозиготных генотипических комбинациях последовательность glaВ2 (glaВ1.2, glaВ2.2) являлась самой распространенной (см. табл. 2, 3). По результатам более ранних исследований инверсионного полиморфизма G. glaucus (г. Калининград, Брянская обл., Саратовская обл.) также был отмечен высокий уровень распространения данных генотипических комбинаций [5, 17].
Относительно низкая частота стандартной последовательности glaВ1 в исследованных водоемах г. Калининграда является, возможно, результатом внутривидовой дивергенции с закреплением гомозиготной инверсии glaВ2. По мнению Беляниной и Дурновой (2006), причиной такой хромосомной дифференциации может быть, к примеру, заселение популяциями одного вида разных водных макрофитов или других субстратов [18]. Поскольку экологические ниши в условиях прибрежной зоны водоемов довольно многообразны, возможно, у хирономид перифитона происходит достаточно быстрое закрепление новой последовательности.
Стоит также отметить последовательности glaВ5 и glaE6 + glaF7, впервые обнаруженные в озере Школьном г. Калининграда (см. табл. 2, 3). В 2010 г. эти последовательности дисков были внесены в реестр последовательностей для G. glaucus г. Калининграда (оз. Пеньковое и система прудов Карасевка). Таким образом, гетерозиготные инверсии В1.5, E1.6, F1.7 продолжают присутствовать и закрепляются в генофонде природных популяций G. glaucus водоемов г. Калининграда [5].
По исследованиям Беляниной (2014), гетерозиготная инверсия D1.3 (0,024) зафиксирована в Брянской обл. (пруд у с. Несвоевка) у одной особи [17]. Гетерозиготная последовательность glaC1.2, обнаруженная в водоемах Брянской и Саратовской областей, имела следующие частоты: 0,02 (Брянская обл., пруд у с. Замишево), 0,25 (Волга у Саратова), 0,44 (пруд у Саратова). Эти последовательности (glaC2 и glaD3) также были обнаружены и в калининградских популяциях G. glaucus (система прудов Карасевка и озеро Школьное) с крайне низкой частотой (от 0,019 до 0,037).
Числа геномных комбинаций и инверсионных последовательностей дисков хромосом G. glaucus водоемов городской черты г. Калининграда показали наибольшие значения данных показателей для озера Школьного (21–11 соответственно) (табл. 4).
Таблица 4. Общие показатели полиморфизма природных популяций Glyptotendipes glaucus водоемов г. КалининградаCommon indicators of polymorphisms of natural populations of Glyptotendipes glaucus in reservoirs of Kaliningrad
Показатели полиморфизма | Карасевка 13.06.12 | Чистый 14.06.12 | Школьное 15.06.12 | Пеньковое 18.06.12 | Мельничный 21.06.12 |
Количество исследуемых особей | 27 | 27 | 52 | 3 | 27 |
Количество особей со стандартным кариотипом | 7 | 13 | 16 | 3 | 15 |
Число полиморфных особей,% | 70,4 | 51,9 | 65,4 | – | 40,7 |
Число гетерозиготных инверсий на особь | 0,93 | 0,63 | 0,75 | – | 0,55 |
Число инверсионных последовательностей дисков | 7 | 5 | 11 | 0 | 3 |
Число геномных комбинаций | 14 | 8 | 21 | 1 | 6 |
Отношение числа инверсионных последовательностей дисков к числу геномных комбинаций (ЧИП/ЧГК) | 0,5 | 0,63 | 0,52 | 0 | 0,5 |
Рассмотренный в более ранних исследованиях показатель отношения числа последовательностей дисков к числу генотипических комбинаций (ЧИП/ЧГК) может служить критерием количественной оценки адаптивных возможностей популяции [5, 19].
Примером применения данного показателя может служить естественная популяция G. glaucus озера Школьного, которое постоянно подвергается воздействию антропогенных бытовых поллютантов и характеризуется как загрязненное (индекс Балушкиной — 6,5) [20]. Подвергающаяся постоянному пролонгированному воздействию поллютантов данная популяция быстрее генерирует большее разнообразие инверсионных последовательностей, при этом число их геномных сочетаний возрастает незначительно, что в итоге приводит к снижению величины этого показателя в условиях возрастающего уровня загрязнения [5, 19].
Таким образом, природная популяция G. glaucus озера Школьного находится в процессе приспособления к новым экологическим условиям, расширяя диапазон хромосомного полиморфизма для вида в целом.
В качестве одного из основных параметров кариологической характеристики популяции используют показатель числа гетерозиготных инверсий, обозначающий уровень естественного хромосомного полиморфизма для каждого вида. Полиморфные виды хирономид, а также других двукрылых насекомых превалируют над мономорфными видами как по территориальному распределению, так и по пластичности приспособления к ареалам с контрастными условиями существования. Так, выраженная дивергенция отдельных популяций хирономид по числу гетерозиготных инверсий указывает на адаптивные возможности вида.
В июне 2012 г. в природных популяциях личинок хирономид G. glaucus исследованных водоемов г. Калининграда число гетерозиготных особей составило 41–70 %, а число гетерозиготных инверсий на особь — 0,55–0,93. Заметно, что уровень хромосомного полиморфизма ниже, чем в более ранних исследованиях популяций G. glaucus водоемов г. Калининграда.
Наблюдения показали, что прибрежные зоны исследованных водоемов г. Калининграда в течение последних лет подвергаются интенсивной высокоэтажной застройке [21]. Застекленные окна зданий в дневное время отражают световые лучи и искусственно освещаются в темное время суток, создавая повышенный уровень светового излучения, привлекают имаго, как световые ловушки, что может являться дополнительным барьером и препятствовать их активному перемещению [22]. Можно предположить, что такой искусственный изолирующий барьер, препятствующий миграционной активности имаго, вероятно, приводит к снижению потока генов и уровня гетерозиготности популяции G. glaucus [23]. Данное предположение требует экспериментального подтверждения.
Также для сравнительного анализа уровня естественного инверсионного полиморфизма G. glaucus для г. Калининграда мы воспользовались известными данными по Саратовской и Брянской областям (табл. 5), которые показали, что уровень инверсионного полиморфизма популяций г. Калининграда G. glaucus в 2012 г. не превысил верхний уровень для популяций Саратовской и Брянской областей [17].
Таблица 5. Сравнительные данные по инверсионному полиморфизму Glyptotendipes glaucus разных областейComparative data on the inversion polymorphism of Glyptotendipes glaucus from different regions
Область, год, авторы исследований | Саратовская область | Брянскаяобласть | Калининградская область | ||
1998 (Белянина, Дурнова) | 2011 (Белянина) | 2011 (Белянина) | 2007 (Шартон) | 2012 (Винокурова, Калинина, Столь) | |
Доля гетерозиготных особей, % | 48–72 | 75–85 | 50–85 | 90,6–100 | 4–70 |
Число гетерози-готных инверсий на особь | 0,70–1,25 | 0,96–1,90 | 0,61–1,07 | 0,69–1,25 | 0,55–0,93 |
ЗАКЛЮЧЕНИЕ
В результате проведенного исследования цитогенетически был идентифицирован и изучен фитофильный вид G. glaucus из пяти водоемов г. Калининграда (пруды Чистый, Мельничный, система прудов Карасевка, озера Пеньковое и Школьное). Для данного вида были отмечены гетерозиготные и гомозиготные инверсии.
Отмечено сохранение тенденции распространения типичных адаптивных гомо- и гетерозиготных инверсий glaВ1.2, glaВ2.2 и закрепление в кариофонде популяций последовательности дисков glaА2 в гетерозиготном состоянии в прудах Карасевка и Чистый. Наличие в популяции системы прудов Карасевка инверсионных последовательностей glaG4 в гомо- и гетерозиготных состояниях и glaD2 в гетерозиготном состоянии в пруду Чистом также было характерно для более ранних исследований G. glaucus из водоемов г. Калининграда [5].
Сравнивая данные по инверсионному полиморфизму личинок G. glaucus г. Калининграда, Саратовской и Брянской областей, можно предложить рабочую гипотезу о прямой зависимости величины хромосомного полиморфизма инверсионного типа от уровня загрязнения водоемов ионами тяжелых металлов (г. Калининград), долгоживущими радионуклидами (Брянская обл.), нитратами и ионами аммиачного азота (Саратовская обл.) [5, 9, 17, 24]. Данная гипотеза не противоречит закономерностям механизмов адаптаций и процессов микроэволюции организмов вследствие давления отбора.
В Калининградской области вид G. glaucus является массовым, и личинки данного вида могут быть рекомендованы в качестве прототипных для цитогенетического мониторинга качества среды обитания [5, 19, 25]. Кроме того, используя вышеприведенные расчеты, индекс ЧИП/ЧГК и анализируя уровень инверсионного полиморфизма популяций G. glaucus, возможно более полно и количественно оценить генотипическую структуру исследуемой популяции.
БЛАГОДАРНОСТИ
Авторы выражают глубокую благодарность за профессиональную и моральную поддержку д-ру биол. наук, вед. науч. сотруднику Н.А. Петровой (ЗИН РАН, Санкт-Петербург), которая поддержала инициативу данного исследования. Особую благодарность выражаем д-ру биол. наук, проф. Н.А. Дурновой (СГМУ им. В.И. Разумовского, Саратов) за консультации и ценные замечания. Мы признательны рецензентам за труд, проделанный над рукописью, критические замечания и полезные дополнения, которые послужили точности передачи смысла работы.
Авторы заявляют об отсутствии конфликта интересов.
About the authors
Natalia V Vinokurova
Immanuel Kant Baltic Federal University, Kaliningrad
Author for correspondence.
Email: NVinokurova@kantiana.ru
PhD in Biological sciences, senior lecturer, School of Life Sciences Russian Federation
Evgeniya A Kalinina
Immanuel Kant Baltic Federal University, Kaliningrad
Email: EAKalinina1@kantiana.ru
Postgraduate, School of Life Sciences Russian Federation
Edgar E Stol’
Immanuel Kant Baltic Federal University, Kaliningrad
Email: ed-wizard@live.ru
Postgraduate, School of Life Sciences Russian Federation
References
- Белоногова Ю.В., Белянина С.И. Динамика морфофункциональных характеристик политенных хромосом личинок хирономид (Chironomus plumosus (L.) под влиянием ионов кадмия // Новые данные по хирономидологии. – Борок: ИБВВ РАН, 2003. – С. 19–22. [Belonogova YuV, Belyanina SI. Dinamika morfofunktsional’nykh kharakteristik politennykh khromosom lichinok khironomid (Chironomus plumosus (L.)) pod vliyaniem ionov kadmiya. In: Novye dannye po khironomidologii. Borok: IBVV RAN; 2003. P. 19-22. (In Russ.)]
- Петрова Н.А., Жиров С.В. Морфология личинки, структура кариотипа и инверсионный полиморфизм хирономиды из Южно-Африканской Республики (Diptera, Chironomidae) // Вавиловский журнал генетики и селекции. – 2010. – Т. 14. – № 1. – C. 70–78. [Petrova NA, Zhirov SV. Larva morphology, karyotype structure, and inversion polymorphism in a chironomid from the republic of South Africa (Diptera, Chironomidae). Vavilovskii zhurnal genetiki i selektsii. 2010;14(1):70-78. (In Russ.)]
- Кикнадзе И.И., Истомина А.Г. Кариотипы и хромосомный полиморфизм сибирских видов хирономид (Diptera, Chironomidae) // Сибирский экологический журнал. – 2000. – Т. 7. – № 4. – C. 445–460. [Kiknadze II, Istomina AG. Kariotipy i khromosomnyi polimorfizm sibirskikh vidov khironomid (Diptera, Chironomidae). Sibirskii ekologicheskii zhurnal. 2000;7(4):445-460. (In Russ.)]
- Petrova NA, Vinokurova NV. Chromosomal variation of populations of Chironomus plumosus Linnaeus (Diptera: Chironomidae) from lakes of Kaliningrad, Russia. Comparative Cytogenetics. 2007;1(1):51-54.
- Шартон А.Ю., Винокурова Н.В., Данилова М.В., и др. Инверсионный полиморфизм Glyptotendipes glaucus Mg. (Diptera: Chironomidae) из водоемов г. Калининграда // Генетика. – 2010. – Т. 46. – № 7. – C. 786–793. [Sharton AY, Vinokurova NV, Danilova MV, et al. Inversion polymorphism of Glyptotendipes glaucus Mg. (Diptera: Chironomidae) from the reservoirs of Kaliningrad. Russ J Genet. 2010;46(7):786-793. (In Russ.)]. doi: 10.1134/S1022795410070021.
- Кикнадзе И.И., Шилова А.И., Керкис И.Е., и др. Кариотипы и морфология личинок трибы Chironomini. — Новосибирск: Наука, 1991. (Kiknadze II, Shilova AI, Kerkis IE, et al. Kariotipy i morfologiya lichinok triby Chironomini. Novosibirsk: Nauka; 1991. (In Russ.)]
- Чубарева Л.А., Петрова Н.А. Методика приготовления цитологических препаратов для кариологического изучения двукрылых насекомых // Новые данные по кариосистематике двукрылых насекомых. — Л.: Изд-во Зоологического института, 1980. [Chubareva LA, Petrova NA. Metodika prigotovleniya tsitologicheskikh preparatov dlya kariologicheskogo izucheniya dvukrylykh nasekomykh. In: Novye dannye po kariosistematike dvukrylykh nasekomykh. Leningrad: Iz-vo Zoologicheskogo instituta; 1980. (In Russ.)]
- Петрова Н.А., Михайлова П.В., Селла Г., и др. Структурно-функциональные изменения политенных хромосом Chironomus riparius из водоемов Италии, загрязненных тяжелыми металлами // Сибирский экологический журнал. – 2000. – № 4. – С. 511–521. [Petrova NA, Mikhaylova PV, Sella G, et al. Structural-functional changes of the polytene chromosomes Chironomus riparius from the Italian water bodies, polluted by heavy metals. Sibirskii ekologicheskii zhurnal. 2000;(4):511-521. (In Russ.)]
- Belyanina SI, Durnova NA. Morphology and chromosomes of phytophilous Glyptotendipes glaucus (Diptera, Chironomidae) from water pools of Saratov district. 2. Karyotypic analysis. Russian Journal of Zoology. 1998;77(2):243-251.
- Петрова Н.А., Винокурова Н.В., Данилова М.В. Некоторые особенности кариотипа популяции Camptochironomus tentans Северо-Западного региона России // Актуальные проблемы экологической физиологии, биохимии и генетики животных. – Саранск: Изд-во Мордовского университета, 2005. – С. 173–175. [Petrova NA, Vinokurova NV, Danilova MV. Nekotorye osobennosti kariotipa populyatsii Camptochironomus tentans severo-zapadnogo regiona Rossii. In: Aktual’nye problemy ekologicheskoi fiziologii, biokhimii i genetiki zhivotnykh. Saransk: Izdatel’stvo Mordovskogo universiteta; 2005. P. 173-175. (In Russ.)]
- Petrova NA, Vinokurova NV, Danilova MV, Maslova VV. Seasonal variability of the karyotype structure of Chironomus plumosus (Diptera, chironomidae) from a biotope of Kaliningrad. Tsitologyia. 2007;49(10):901-905.
- Kiknadze II, Golygina VV, Broshkov AD, et al. Mystery of Chironomus dorsalis Meigen karyotype (Diptera: Chironomidae). Comparative Cytogenetics. 2008;2(1):21-35.
- 13.Гланц С. Медико-биологическая статистика / Пер. с англ. – М.: Практика, 1998. [Glantz S. Primer of biostatistics. Moscow: Praktika; 1998. (In Russ.)]
- Кикнадзе И.И., Истомина А.Г., Гундерина Л.И., и др. Кариофонды хирономид криолитозоны Якутии. Триба Chironomini. – Новосибирск: Наука, 1996. [Kiknadze II, Istomina AG, Gunderina LI, et al. Kariofondy khironomid kriolitozony Yakutii. Triba Chironomini. Novosibirsk: Science; 1996. (In Russ.)]
- Провиз В.И., Базова Н.В. Кариотип Glyptotendipes glaucus (Diptera, Chironomidae) из озера Котокельское (Прибайкалье) // Зоологический журнал. – 2012. – Т. 91. – № 2. – С. 247–251. [Proviz VI, Bazova NV. Karyotype Glyptotendipes glaucus (Diptera, Chironomidae) from the lake Kotokelskoe (Baikal). Zoologicheskii Zhurnal. 2012;91(2):247-251. (In Russ.)]. doi: 10.1134/S0013873812030128.
- Ilkova J. Chromosome variability of species of subfamily Chironominae (Diptera, Chironomidae) from polluted regions near Sofia. Acta Zoologica Bulgarica. 2004;56(1):15-30.
- Белянина С.И. Сравнительный хромосомный анализ популяций фитофильной хирономиды Glyptotendipes glaucus (Mg.) из водоемов, затронутых чернобыльским выбросом // Генетика. – 2014. – Т. 50. – № 9. – С. 1025–1032. [Belyanina SI. Comparative chromosomal analysis of populations of phytophilous chironomidae Glyptotendipes glaucus (Mg.) from Chernobyl-Affected Territory. Russ J Genet. 2014;50(9):1025-1032. (In Russ.)]. doi: 10.1134/S1022795414090038.
- Дурнова Н.А., Белянина С.И. Экологическая и хромосомная эволюция хирономид перифитона (Diptera, Chironomidae) / IV Международная конференция по кариосистематике беспозвоночных животных; 28–30 августа 2006 года; Санкт-Петербург. [Durnova NA, Belyanina SI. Ekologicheskaya i khromosomnaya evolyutsiya khironomid perifitona (Diptera: Chironomidae). (Conference proceedings). IV Mezhdunarodnaya konferentsiya po kariosistematike bespozvonochnykh zhivotnykh; 2006 aug 28-30; Saint Petersburg. (In Russ.)]
- Винокурова Н.В., Шмаров В.А. Оценка влияния половой дифференциации, сезона генерации и тяжелых металлов на параметры хромосомного инверсионного полиморфизма Glyptotendipes glaucus Mg. (Diptera: Chironomidae) / V Международная заочная научно-практическая конференция «Тенденции развития естественных и математических наук»; 28 апреля 2013 года; Новосибирск. [Vinokurova NV, Shmarov VA. Otsenka vliyaniya polovoi differentsiatsii, sezona generatsii i tyazhelykh metallov na parametry khromosomnogo inversionnogo polimorfizma Glyptotendipes glaucus Mg. (Diptera: Chironomidae). (Conference proceedings). V Mezhdunarodnaya zaochnaya nauchno-prakticheskaya konferentsiya “Tendentsii razvitiya estestvennykh i matematicheskikh nauk”; 2013 apr 28; Novosibirsk. (In Russ.)]
- Гапаненок Е.А., Винокурова Н.В. Сравнительные данные по фауне хирономид пруда Школьного города Калининграда / XXVII Международная научно-практическая конференция «Естественные и математические науки в современном мире»; 4 февраля 2015 года; Новосибирск. [Gapanenok EA, Vinokurova NV. (Conference proceedings). XXVII Mezhdunarodnaya nauchno-prakticheskaya konferentsiya “Estestvennye i matematicheskie nauki v sovremennom mire”; 2015 feb 04; Novosibirsk. (In Russ.)]
- Правительство Калининградской области. Об экологической обстановке в Калининградской области в 2015 году. – Калининград: Государственный доклад, 2016. – 200 с. [Pravitel’stvo Kaliningradskoi oblasti. Ob ekologicheskoi obstanovke v Kaliningradskoi oblasti v 2015 godu. Kaliningrad: Gosudarstvennyi doklad; 2016. 200 p. (In Russ.)]
- Hirabayashi K. Studies on the Massive Flights of Chironomid Midges (Diptera: Chironomidae) as Nuisance Insects and Plans for their Control in the Lake Suwa Area, Central Japan 1. Occurrence of massive flights of Tokunagayusurika akamusi. Nihon Eiseigaku Zasshi. Japanese Journal of Hygiene. 1991;46(2):652-661. doi: 10.1265/jjh.46.652.
- Алтухов Ю.П., Салменкова Е.А., Курбатова О.Л., и др. Динамика популяционных генофондов при антропогенных воздействиях. – М.: Наука, 2004. [Altukhov YP, Salmenkova EA, Kurbatova OL, et al. Dinamika populyatsionnykh genofondov pri antropogennykh vozdeistviyakh. Moscow: Nauka; 2004. (In Russ.)]
- Столь Э.Э., Калинина Е.А. Свинец и цинк в донных отложениях водоемов Калининградской области / Всероссийская (с международным участием) научная школа-конференция, посвященная 115-летию со дня рождения А.А. Уранова. «Cовременные концепции экологии биосистем и их роль в решении проблем сохранения природы и природопользования»; 10–14 мая 2016 года; Пенза. [Stol’ EE, Kalinina EA. Svinets i tsink v donnykh otlozheniyakh vodoemov Kaliningradskoi oblasti. (Conference proceedings). Vserossiiskaya (s mezhdunarodnym uchastiem) nauchnaya shkola-konferentsiya, posvyashchennaya 115-letiyu so dnya rozhdeniya A.A. Uranova. “Covremennye kontseptsii ekologii biosistem i ikh rol’ v reshenii problem sokhraneniya prirody i prirodopol’zovaniya”; 2016 May 10-14; Penza. (In Russ.)]
- Винокурова Н.В., Шартон А.Ю. Региональные кариотипические особенности Glyptotendipes glaucus (Diptera: Chironomidae) из водоемов города Калининграда / Сборник тезисов X съезда Гидробиологического общества при РАН; Владивосток, 28 сентября — 2 октября 2009 года. – Владивосток: Дальнаука, 2009. – С. 72. [Vinokurova NV, Sharton AY. Regional’nye kariotipicheskie osobennosti Glyptotendipes glaucus (Diptera: Chironomidae) iz vodoemov goroda Kaliningrada. In: Proceedings of the X S»ezda Gidrobiologicheskogo obshchestva pri RAN; Vladivostok, 28 September – 2 October 2009. Vladivostok: Dal’nauka; 2009. P. 72. (In Russ.)]
Supplementary files


