







О биологическом и социальном в сексуальной ориентации человека: хемокоммуникационная гипотеза

- Авторы: Даев Е.В.1
-
Учреждения:
- ФГБУ ВПО «Санкт-Петербургский государственный университет»
- Выпуск: Том 14, № 3 (2016)
- Страницы: 3-12
- Раздел: Статьи
- Статья получена: 14.11.2016
- Статья опубликована: 15.09.2016
- URL: https://journals.eco-vector.com/ecolgenet/article/view/5478
- DOI: https://doi.org/10.17816/ecogen1433-12
- ID: 5478

Цитировать
Полный текст

Аннотация
Недопонимание роли биологических и социальных факторов в формировании некоторых социально значимых признаков у человека могут приводить к возникновению неоправданной напряженности в межличностных отношениях. Это связано с частой недостоверностью информации, ее искаженным восприятием, размытостью из-за нечеткости используемой терминологии и, как следствие, невозможности ее корректного анализа воспринимающим человеком. На примере признака «сексуальная ориентация» показано, как генетические представления о формировании признака и данные о механизмах контроля формирования пола могут уточнить и дополнить наши знания по этому вопросу. В рамках темы рассмотрена гипотеза о хемокоммуникационной модели формирования «сексуальной ориентации» у человека.
Ключевые слова
Полный текст
ВВЕДЕНИЕ
Синэкологические внутривидовые взаимоотношения, в особенности регулирующие репродукцию вида, могут играть существенную роль в формировании наследственной изменчивости и пространственно-временной структуры популяций животных [1, 2]. Тем не менее научному изучению этих аспектов межорганизменных взаимоотношений, особенно у человека, уделяется недостаточно внимания. Представляется перспективным с этой точки зрения рассмотреть возможные механизмы формирования сексуальной ориентации человека как одного из факторов, определяющих репродуктивный потенциал вида Homo sapiens.
В настоящее время проблема существования различных вариаций сексуальной ориентации (далее — СОя) привлекает все большее внимание. Ввиду неоднозначного отношения мирового сообщества к некоторым вариантам СОи, вопрос о соотношении биологических, генетических и социальных механизмов ее формирования приобретает особую актуальность [3].
Сравнение публикационной активности по данным базы Scopus за каждые 5 лет с 1996 по 2015 год показывает:
а) при росте числа публикаций по вопросам полового поведения человека (human sexual behavior) в 2,3 раза (с 10 422 в первые пять лет до 24 185 за последние пять лет) доля в них статей, содержащих термины «ген» и «генетический», возросла с 5 до 8 %;
б) наоборот, при постоянном росте числа научных работ, посвященных сексуальной ориентации (четырехкратный рост при сравнении первой и последней пятилеток), доля в них работ с генетической терминологией снизилась с 8–10 до 5 %.
Одна из возможных причин снижения частоты публикаций с «генетическим» уклоном о проблемах СОи может крыться в нечеткости определения изучаемого признака и его (определения) несоответствии формальным требованиям классической генетики.
Общие представления о признаке «сексуальная ориентация»
Сексуальная ориентация — это признак, выражающийся в половом влечении к людям своего пола (гомосексуальность), противоположного пола (гетеросексуальность) или обоим полам (бисексуальность) [4, 5]. Ее определяют так же, как направленность полового интереса к лицам своего, противоположного или к лицам обоих полов, особенно когда эта направленность продиктована скорее физиологическими, чем социологическими причинами [6]. Можно найти определение СОи как варианта предпочтения сексуальной активности с лицами определенного пола [6] и др. Сразу заметна неоднозначность таких определений из-за использования крайне расплывчатых по содержанию слов типа: «влечение», «интерес», «активность» и т. д. Так, например, сексуальное влечение (attraction) разделяют на физическое и романтическое или эмоциональное [4]. Подобные эпитеты мало что прибавляют к научным знаниям о сути признака «СОя».
По своей природе признак «СОя» крайне сложен (complex trait) и состоит из множества лабильных компонентов, в каждом конкретном случае зависящих от взаимодействия уникального генотипа с не менее уникальными сочетаниями постоянно меняющихся факторов окружающей среды.
Чтобы лучше разобраться в том, что представляет собой СОя как фенотипический признак для последующего научного изучения, следует всегда помнить, что в каждом конкретном случае так называют специфическую совокупность форм поведения изучаемого человека по отношению к другим людям. В качестве рабочего определения назовем СОю результирующим поведенческим ответом на анализ и последующий синтез мозгом человека всей поступающей извне информации о первичных, вторичных и третичных половых признаках окружающих людей. Причем уникальность СОи каждого человека детерминирована биологическими и гендерными особенностями состояния его организма в каждый момент времени. Только по специфике проявления в виде какой-либо формы (нескольких форм) поведения исследователь может оценить СОю. Тем не менее этот «суммирующий» комплекс поведенческих признаков — важнейший фактор синэкологических взаимоотношений, непосредственно связанный с репродукцией как у человека, так и у других видов животных, размножающихся половым путем.
Попробуем в упрощенном варианте представить себе последовательность событий, которая приводит в конечном счете к формированию СОи. Для начала попробуем дать рабочее определение признака «пол», потому что сексуальная ориентация неразрывно связана с существованием мужского и женского пола и их поведенческими различиями.
О дифференцировке по полу и половому поведению
Термин «пол» можно определить следующим образом.
Пол — это наследуемый как единое целое многокомпонентный комплекс специфических характеристик, проявляющийся в виде двух фенотипов, которые у человека обозначены как «женский» и «мужской» («самец» и «самка» у животных).
На самом деле мужской и женский пол представляют собой два множества фенотипов, которые различают по набору специфических характеристик. Иногда целостность такого набора может быть существенно нарушена, что не позволяет точно определить пол. В этом случае особь (животное или человек) называют интерсексом.
Согласно одной из предлагаемых моделей половой дифференцировки у млекопитающих на начальном этапе все зависит от генов половых хромосом. Они контролируют не только возникновение гонад, но и участвуют в локальной специализации тканей мозга посредством взаимодействия гормональных и автономных клеточных механизмов с пока не идентифицированными регуляторными молекулами [7, 8].
Принято считать, что пол у человека определяется в первую очередь составом половых хромосом (XX или XY), попавших в зиготу. Согласно 83-му выпуску Ensembl genome browser release 2015 года [9] длина Y-хромосомы человека, генетический материал которой лежит в основе различий между женским и мужским фенотипами, представляет всего лишь ≈1,6 % референсного генома (≈57/3547 Mb). Учитывая диплоидность, различия между женским и мужским организмами по количеству генетического материала составляют всего ≈0,8 % и даже еще меньше, если учитывать наличие псевдоаутосомных районов. По числу кодирующих генов разница составляет ≈0,2–0,4 %. Таким образом, мы, вероятно, сильно преувеличиваем и абсолютизируем разницу между женским и мужским фенотипами. Исходя из упрощенного представления о пропорциональности количества генов числу фенотипических признаков, которые контролируются этими генами, различие должно составлять всего 0,2–0,4 % от всех фенотипических признаков человека (рис. 1).
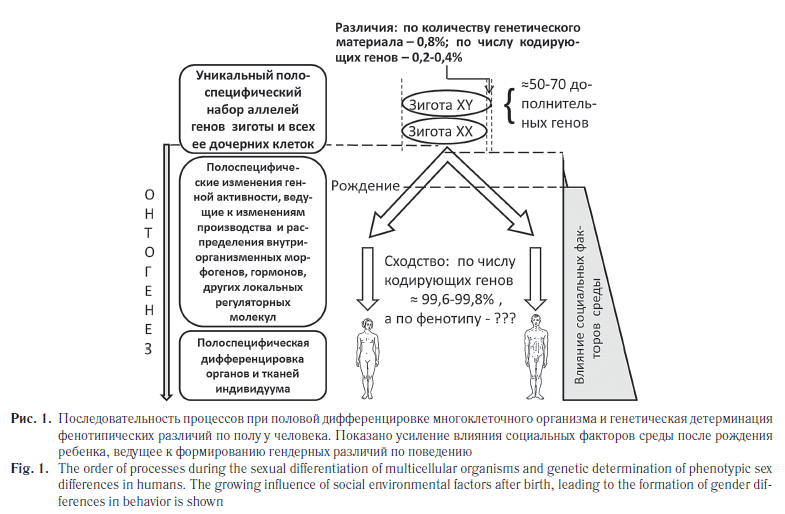
В то же время данные исследований показывают, что мужской и женский фенотипы человека зависят и от аллельного состава родительских гаплотипов аутосом. Достаточно, например, встретиться в зиготе двум рецессивным мутантным аллелям гена CYP21A2 (6-я хромосома), кодирующего фермент стероидогенеза в надпочечниках — 21-гидроксилазу, как организм с женскими половыми хромосомами фенотипически может превратиться в мужской. Степень маскулинизации будет зависеть от сочетания конкретных мутантных аллелей [10]. В крайних вариантах мужчину с ХХ-хромосомами можно будет отличить от мужчины с XY-хромосомами, только если он обратится к медикам по вопросу причин бесплодия в браке.
Во 2-й аутосоме расположен ген SRD5A2. Аллели этого гена кодируют разные варианты фермента 5-альфа-редуктазы, конвертирующей тестостерон в дегидротестостерон. Мутации, нарушающие активность фермента, вызывают более или менее выраженную феминизацию у людей с XY-хромосомами [11].
К феминизации мужского и женского фенотипа ведет наличие некоторых аллелей гена ароматазы (CYP19A1), локализованного в 15-й хромосоме человека [12].
Вполне вероятно, что нами существенно недооценен вклад аутосомных генов, участвующих в детерминации пола у человека (кроме CYP21A2, SRD5A2 и некоторых других). Это подтверждается на модельном объекте — трансгенных мышах XEOSox9 и XOSry, Eif2s3x. В экспериментах по трансгенной гиперактивации гена Eif2s3x Х-хромосомы и аутосомного гена Sox9показано, что индуцировать сперматогенез и получать потомство от мышей можно в отсутствие Y-хромосомы, хотя именно ею в естественных условиях определяется фенотип «самца» [13].
Таким образом, между типичными женским и мужским фенотипами может возникать множество «интерсексных» вариантов с разной степенью выраженности того или иного фенотипа и разными вариантами СОи. Все зависит от конкретной мутации в том или ином гене, состава аллелей по другим генам, условий окружающей среды, включающей в том числе и социально обусловленные факторы.
Самый известный на данный момент синдром нечувствительности к андрогенам (AIS) связан с мутациями в гене AR (на X-хромосоме), кодирующем рецептор андрогенов. В некоторых случаях человек с XY-половыми хромосомами может быть неотличим по фенотипу (за исключением бесплодия) от женщины с двумя Х-хромосомами. При этом в случае «крайних вариантов» фенотипических изменений в мужскую или женскую сторону СОя часто типична для соответствующего фенотипа, а у «интерсексов» возникают определенные проблемы.
У людей с типичным мужским или женским фенотипом, соответствующим набору половых хромосом, может формироваться гомосексуальная ориентация. Методом полногеномного анализа недавно показано, что по крайней мере у мужчин формирование СОи сцеплено с перицентромерным районом q11.23 на 8-й хромосоме и районом q28 Х-хромосомы. В зоне 8q11.23 расположен ген рецептора нейропептида B/W, представленного на мембранах клеток лимбической системы мозга и гиппокампа. Взаимодействие этого рецептора с лигандом модулирует социальные взаимодействия у мышей, а у человека изменяет эмоциональный ответ на выражение лица [14]. В районе Xq28 находится ген одного из рецепторов аргинин-вазопрессина — гормона, модулирующего социальное поведение. В этом же районе расположен еще один ген, экспрессирующийся в мозге. Это ген CNGA2 (цАМФ-зависимого ионного канала α-2), аллели которого регулируют формы социального и полового поведения у мышей, индуцированные запахами, в том числе зависимыми от гаплотипа главного локуса гистосовместимости [14]. Как показывают исследования, СОя женщин также зависит от влияния не только средовых, но и генетических факторов [15].
Взаимодействие уникального наследственного материала аутосом и половых хромосом с еще более уникальным сочетанием средовых факторов в процессе онтогенетического развития только усиливает генетическую и фенотипическую изменчивость по половым признакам по мере усложнения формирующегося многоклеточного организма. Сюда относится и изменчивость форм поведения, в том числе сцепленных с полом, ограниченных полом или зависимых от пола.
Наиболее полноценно поведение человека можно изучать, начиная с постнатальных стадий его онтогенетического развития. По некоторым данным, различия в поведении у девочек и мальчиков можно регистрировать уже на первый день после рождения и определенно в возрасте 3–12 месяцев. Это касается таких признаков, как фиксация взгляда на определенных предметах, темперамент, предпочтение определенных игрушек и игр и т. д. [4, 16–18]. Их надо четко отличать от специфических форм полового поведения (ПП). Последние, видимо, возникают на основе исследовательского поведения, когда ребенок начинает осознавать существование половых различий и идентифицировать свою принадлежность к определенному полу.
Очевидно, что различия в ПП между мальчиками и девочками проявляются намного позже, чем, например, биохимические, морфологические и целый ряд других признаков (в том числе поведенческих). Можно полагать, подобные различия (зависимые от пола ребенка) происходят из врожденной и/или условно-рефлекторно возникающей привлекательности/непривлекательности отдельных свойств (характеристик, черт) окружающих людей, когда исследовательское поведение концентрируется на комплексе признаков, связанных с тем или иным полом. Формирование у реципиента полоспецифических сигналов является результатом их анализа мозгом, точность работы которого зависит в том числе и от работы всех аллельных вариантов генов конкретного индивида. Так как с момента рождения резко усиливается влияние социально обусловленных факторов окружающей среды на организм ребенка (см. рис. 1), их разнообразие и сочетанное действие на конкретный генотип, особенно в критические периоды постнатального онтогенеза, могут приводить к формированию уникального варианта СОи.
Различия в ПП мальчиков и девочек и, собственно, СОя контролируются мозгом, и их можно рассматривать как вторичные характеристики, которые зависят от специфики межнейронных связей, объема специфических ядер головного мозга, синаптической плотности на дендритах нейронов [4, 19]. В свою очередь морфологические особенности строения мозга закладываются на относительно коротком этапе развития под влиянием гормонов (эстрадиола, простагландинов и др.) [17], а следовательно, и аллелей генов, контролирующих различные этапы синтеза этих гормонов. Слабые, пока не исследованные отклонения эндокринного баланса в раннем онтогенезе могут оказывать разнонаправленное влияние на СОю и другие проявления ПП у взрослых организмов, многократно усиливая индивидуальную изменчивость [19, 20].
СОя как многокомпонентный признак
А. Кинси, анализируя множество индивидуальных случаев, подчеркивал, что существует «непрерывность градаций» от «исключительно» гетеросексуальных до «исключительно» гомосексуальных индивидуумов. «Но огромное количество людей не хотят верить в это. Они пытаются подойти к вопросу (как и ко многим другим. — Прим. авт.) дихотомически, деля половое поведение на „нормальное“ и „аномальное“, социально приемлемое и не приемлемое, гетеросексуальное и гомосексуальное» [21].
Многие исследователи понимают необходимость введения точного определения признака СОя, от чего зависит выбор пути его изучения. В настоящее время сексуальную ориентацию принято оценивать по трем параметрам: «сексуальному влечению», «сексуальному поведению» и «самоидентификации» [22, 23]. Однако почему-то почти не обсуждается ни сложность объективной оценки этих многофакторных параметров, ни взаимозависимость этих параметров друг от друга. Так, основным компонентом признака СОя считается сексуальное влечение (attraction) к лицам определенного пола. Сексуальное (половое) поведение — это проявление (манифестация) СОи в отношении притягательного объекта. Само-идентификацию же некоторые авторы рассматривают только как оценку человеком собственной СОи: гомо-, би- или гетеросексуальной. Между тем очевидно существование гораздо большего числа градаций, чем могут предложить исследователи, поэтому они вводят все большее число измеряемых параметров, по которым можно точнее оценить гетерогенность Homo sapiens по признаку СОи [22].
Основным компонентом СОи можно дать разные определения. Так, под само-идентификацией, например, можно понимать самоощущение человеком собственного пола: мужского или женского. Манифестация «сексуального влечения» может заключаться в совершенно разных формах поведения, не обязательно полового. Все это добавляет сложности к изучению проблемы формирования различных форм СОи.
Рассматривая историю вопроса о COи в разных культурах, Ли Вэй отмечает большую роль ограниченности выбора (распространение однополых изолятов с социально ограниченной возможностью контактов между полами) в формировании гомосексуального поведения [4]. Негативный опыт (физическое, сексуальное, эмоциональное насилие, бытовые злоупотребления и др.), приобретенный в детстве, повышает вероятность возникновения «нетрадиционной» СОи [24, 25]. В свою очередь в группах сексуальных меньшинств выявлен высокий риск возникновения умственных расстройств [26]. Причем устойчивое словосочетание «нетрадиционная COя» только подчеркивает социальный компонент в ее формировании.
Морально-этические нормы ограничивают возможности изучения формирования СОи у человека. Чаще всего ее анализируют косвенно (по самооценке респондентов при анкетировании), констатируя частоту встречаемости в выборках нескольких типов СОи (в основном гетеро-, би- и гомосексуальности). Такая оценка может быть существенно искажена по целому ряду причин социального и личного характера участников анкетирования из-за чего возможная роль наследственного материала маскируется влиянием социальной среды. Тем не менее сравнение частот в выборках, сформированных по разным принципам, выявляет интересные закономерности, например, связь гомо- и бисексуальности с насилием (физическим, психическим или эмоциональным) в детском возрасте [24, 25]. Гораздо реже для оценки СОи используют более прямые измерения (разными методами) сексуального возбуждения испытуемых при показе, например, фотографий с эротическим содержанием [4].
Анализ современной литературы приводит к мысли, что все варианты сексуальной ориентации формируются работой единого механизма контроля ПП, возникшего в процессе эволюции. При этом сексуальная привлекательность обусловлена взаимодействием генетических, онтогенетических, культурных и психологических факторов [27]. Так как признак СОи имеет ярко выраженную социальную окраску, роль генетической составляющей ни в коем случае нельзя ни преуменьшать, ни преувеличивать. Неправильное представление о роли генов в формировании этого признака может существенно влиять на поведение общества и принятие им правовых и политических решений [3].
С генетической точки зрения представляется целесообразным в проблеме СОи выделить четыре основных вопроса, представляющих интерес для генетиков:
1) какой генетический материал достался организму от родителей (генеалогический подход);
2) как происходит половая дифференцировка в пре- и постнатальном онтогенезе на уровне регуляции генной активности клеток различными внешними факторами;
3) как различные наследуемые аномалии формирования пола и ряда других признаков могут быть связаны с различными вариантами СОи, т. е. механизмы изменчивости и наследования последних;
4) какие генетические следствия для популяций человека может иметь наличие различных СОи.
Из вышеперечисленных направлений первые два изучают: а) возможные предпосылки и б) механизмы формирования различных вариантов половой дифференцировки — той биологической основы, на которой формируется СОя.
В последних двух направлениях (касающихся непосредственно СОи) нужно учитывать не только биологические факторы, но и влияние гендерного воспитания, а также других социально обусловленных факторов. Изучать сложный поведенческий признак СОи следует пошагово, предварительно раздробив его на как можно большее число по возможности «элементарных» компонентов. Перспективным представляется также поиск факторов, модулирующих эти компоненты на разных этапах формирования СОи. Чем проще модулятор, тем яснее природа механизма, формирующего и изменяющего признак.
Чем элементарнее анализируемый признак, тем более однозначно он определяем. В вопросе изучения СОи остро назрела проблема унификации используемых терминов. Нужно правильно распознавать, когда исследователи используют одну и ту же терминологию для описания совершенно разных явлений. Не менее трудно сопоставлять случаи, когда экспериментаторы изучают одно и то же явление, но используют разную терминологию. Примером является описание процесса выбора полового партнера [28], который можно рассматривать как одно из проявлений СОи.
Особое внимание следует обратить на консерватизм генетических механизмов формирования пола и зависимых от него форм ПП, что позволяет использовать модельные объекты, в том числе и для изучения формирования СОи. Далее, я попробую вычленить только один из эволюционно древних механизмов, модулирующих ПП животных, и попытаюсь рассмотреть его возможный вклад в изменчивость СОи у человека.
Вклад хемосигналов в формирование сексуальной ориентации
Хемокоммуникационная модель
Коль [29] отмечает, что из-за сильного влияния социальных факторов, таких как гендерное воспитание, биологические основы формирования сексуальных предпочтений у человека могут легко остаться незамеченными. Между тем они могут быть результатом неосознаваемых изменений гормонального статуса организма вследствие еще более трудно осознаваемых влияний полоспецифических химических сигналов (предположительно феромонов), начинающих действовать на самых ранних этапах онтогенеза. Кроме того, могут возникать условно-рефлекторные связи между ольфакторными и «неольфакторными» социально обусловленными факторами. Показано, что нос человека сохраняет высокую чувствительность по крайней мере к некоторым летучим соединениям, действие которых на организм может не осознаваться. Хемосигналы могут менять поведение, настроение, физиологические и когнитивные процессы, действуя на уровне подсознания [30].
О роли феромонов в регуляции ПП говорят исследования на разных видах животных [31]. В свете данных о некоторых эффектах, выявленных у человека [32–34], вполне вероятно, что и такой признак, как Соя, может зависеть от летучих хемосигналов. Связь генетически детерминированных дефектов обоняния (аллели генов ANOS1, FGFR1, PROK2 и PROKR2, локализованных в 10, 8, 3 и 20-й хромосомах соответственно, и др.) с нарушением проявления репродуктивно важных признаков человека (синдром Каллманна) может быть обусловлена изменениями или отсутствием регулирующего влияния хемосигналов, действующих через систему органов обоняния [35, 36]. Тем не менее возможный вклад хемокоммуникационного механизма в формирование ПП и СОиостается практически не исследованным.
Наши наблюдения на модельном объекте — лабораторных мышах (не опубликовано) показывают, что их «сексуальная ориентация», оцениваемая по проявлениям полового поведения (ухаживание, попытки копуляции и т. п.), детерминирована хемосигналами мочи. Нанесение свежесобранной мочи самца на самку сразу меняет поведение тестерного самца с полового на агрессивное. В то же время нанесение мочи самки на самца также меняет поведение самца-тестера на противоположное: вместо агрессии начинают проявляться формы полового поведения (ухаживание, попытки скрещивания). Таким образом, по отношению к самке, пахнущей самцом, как и к самцу, пахнущему самкой, мы наблюдаем инверсию форм полового поведения, которое можно трактовать как гомосексуальное.
Принято считать, что у человека взаимоотношения между людьми регулируются в первую очередь через языковое общение (слух) и зрительное восприятие. Однако хемокоммуникация посредством феромонов может играть не менее важную роль в формировании симпатий-антипатий, половых предпочтений и других форм поведения. В пользу этого свидетельствуют данные работы Martins et al. [37], которые показали, что люди дают самую негативную оценку запаху гомосексуально ориентированных мужчин. Последние же относятся к нему наименее негативно. Это подтверждает существование биохимических различий в составе феромонов и/или в их рецепции между гомо- и гетеросексуально ориентированными людьми.
Если предположить, что хемосигналы человека играют роль в формировании его СОи, а их синтез и рецепция находятся под генетическим контролем, то можно создать упрощенную модель, частично объясняющую существование различных вариантов этого признака [38]. Она является дальнейшим развитием представлений Коля о пятиэтапном становлении СОи у человека от клеточного к организменному уровню [29].
Достаточно представить, что каждый человек является источником как «женских», так и «мужских» аттрактивных (привлекающих) хемосигналов, которые синтезируются из общих предшественников. Следует также допустить, что к своим собственным полоспецифическим хемосигналам развивается относительная нечувствительность (наподобие толерантности к собственным антигенам). Такое допущение вытекает из общности защитных функций иммунной системы и ольфакторного механизма детекции потенциально полезных или вредных факторов окружающей среды. Кроме того, возможно взаимодействие механизмов поиска здоровых и наиболее подходящих по HLA-статусу половых партнеров с механизмами модуляции иммунного ответа [39]. Может также существовать дозовая зависимость чувствительности/нечувствительности к производимым хемосигналам, что косвенно подтверждается данными некоторых исследований [13, 40].
Баланс продукции «мужских» и «женских» хемосигналов зависит от регулирующего действия в первую очередь пары половых хромосом. В наиболее типичном случае у женщин «женские» хемосигналы преобладают над мужскими — результат работы XX-хромосом, а у мужчин — наоборот (рис. 2). В этом случае преобладающий аттрактивный эффект «мужских» феромонов, к которым не была выработана «толерантность», определяет гетеросексуальную ориентацию женщин (см. рис. 2, а).
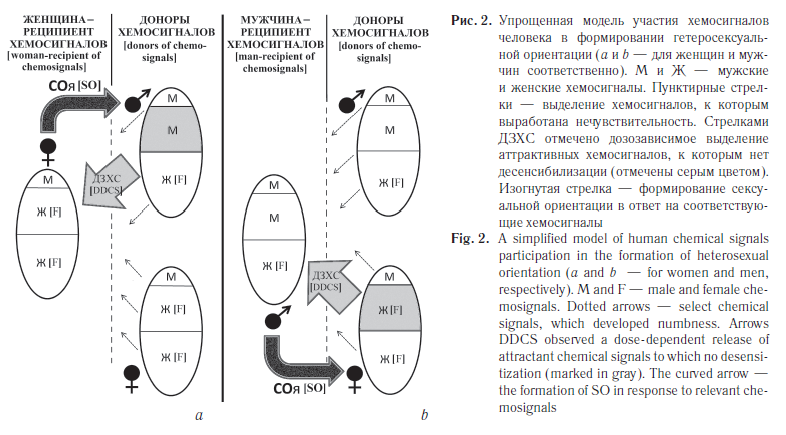
У мужчин, которые нечувствительны к собственным сигналам — «мужским» и «женским», последних меньше, чем у женщин, что определяется в первую очередь регуляторным влиянием пока не установленных генов Y-хромосомы. Поэтому основным фактором формирования их (мужчин) гетеросексуальной ориентации будет качественное и количественное преобладание «женских» хемосигналов у женщин. Оно возникает из-за гетерозиготности по ряду генов Х-хромосом, независимой «лайонизации» и отсутствия Y-хромосомного контроля (см. рис. 2, b).
То, что у женщин и мужчин в течение всей жизни не вырабатывается нечувствительность к «чужим» полоспецифическим сигналам, свидетельствует о существовании критического периода в раннем онтогенезе человека, когда формируется «толерантность» к собственным сигналам (т. е. отсутствие на них выраженной реакции).
Что же касается биологических предпосылок возникновения гомосексуальных вариантов ориентации, то они могут заключаться в специфических дефектах рецепции хемосигналов у отдельных людей — «реципиентов ХС» (рис. 3) или в дефектах производства людьми — «донорами ХС» полоспецифических хемосигналов (рис. 4). Правда, в последнем случае речь пойдет не об особых вариантах СОи, а скорее о несоответствии хемокоммуникационной информации полу конкретного донора хемосигналов.
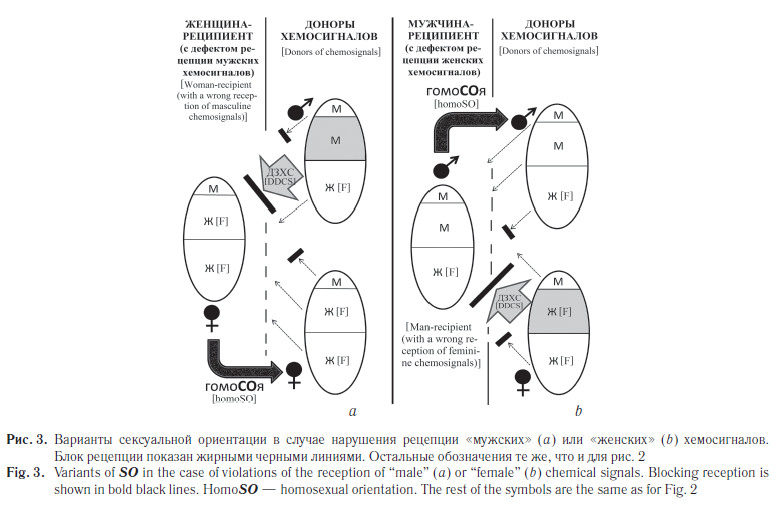
Так, если в организме женщины полностью нарушена рецепция «мужских» хемосигналов, то аттрактивность мужчин (одна Х-хромосома) для этой женщины будет ниже, чем женщин, которые производят больше «привлекающих» хемосигналов вследствие гетерозиготности и/или дозовой компенсации (две Х-хромосомы). Этот фон может быть причиной возникновения гомосексуальной ориентации (см. рис. 3, а). На ориентацию мужчины такой дефект влияния не окажет. Точно так же дефект рецепции «женских» хемосигналов не окажет влияния на СОюженщины, а вот у мужчины может возникать влечение к особям своего пола (см. рис. 3, b).
Помимо дефектов рецепции полоспецифических хемосигналов в организме человека могут возникать различные нарушения их синтеза. В этом случае нарушение баланса «мужских» и «женских» хемосигналов может вести к изменению ориентации в отношении него со стороны окружающих (см. рис. 4). Нарушение производства «мужских» сигналов в мужском организме и «женских» в женском будет приводить к возникновению гомосексуального интереса к таким людям.
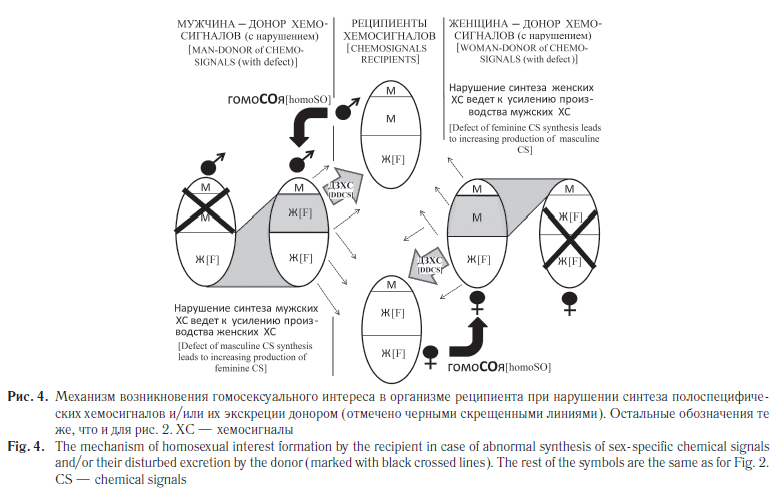
Предлагаемая упрощенная схема, конечно, не охватывает всего многообразия вариантов генетических и иных нарушений, которые могут вести к функциональным нарушениям разной степени выраженности. На них накладываются особенности внутриутробной стадии развития, действие фармакологических препаратов, воспитание в детский период, возникающие условно-рефлекторные связи и импринтинг, та или иная гендерная информация, социальные факторы, действующие в критические периоды развития, культурные и другие особенности [4, 41]. Это неизбежно приводит к многообразию состава компонентов, которые формируют СОю, проявляющуюся в виде различных форм полового и неполового поведения. Исследования на животных показывают, что состав хемокоммуникационных сигналов меняется в зависимости от плотности содержания, микрофлоры, инфекций и воспалительных процессов, протекающих в организме, мозговых травм [42–44]. Как следствие, можно полагать, что признак СОя (по крайней мере, степень его выраженности) тоже модулируется физиологическим состоянием как донорского, так и реципиентного организма.
В каждом конкретном случае вероятность формирования СОи может зависеть от множества генетически детерминированных морфологических, биохимических и других характеристик, которые могут не соответствовать набору половых хромосом. Нарушение работы эволюционно консервативного механизма хемокоммуникации также может существенно влиять на СОю. Действие социальных факторов окружающей среды существенно усугубляет или сглаживает это влияние. На основе данных литературы [24, 25] можно полагать, что неблагоприятное воздействие социальной среды в критические периоды развития (искусственно создаваемая сенсорная депривация при перенасыщенности социальной информацией, ошибки гендерного воспитания и т. п.) может само по себе формировать ту или иную СОю.
Таким образом, вопрос о степени влияния генетических или социальных факторов на СОю имеет смысл только для каждого конкретного случая в каждый момент времени: он может оказаться бесконечно малым или стремиться к единице в зависимости от уникального сочетания всех действующих факторов. Что же сильнее повлияло, гены или среда, никоим образом не должно влиять на специфику отношения общества к любому варианту СОи, если это не идет вразрез с общемировыми нормами морали, этики и в первую очередь уголовного и гражданского права.
СОя играет важную роль как индикатор индивидуального здоровья и социальной изоляции [22]. Для генетики особый интерес представляют поиск и изучение тех случаев, когда можно выявить существенное влияние генетических факторов на формирование отдельных компонентов СОи в конкретных средовых условиях и степень ее выраженности. Последнее, в свете предложенной хемокоммуникационной гипотезы, может объяснять описанное исследователями явление ассортативности заключения браков у человека, ведущее к увеличению генетического разнообразия в потомстве через восприятие запаха гаплотипов главного комплекса гистосовместимости [45]. Перспективным направлением исследований кажется поиск генетических детерминант синтеза и экскреции полоспецифических хемосигналов (феромонов), которые как в пре-, так и в постнатальном периоде онтогенеза (подсознательно) могут вносить свой вклад в формирование сексуальной ориентации человека.
Работа поддержана грантом Президента РФ по поддержке ведущих научных школ и грантом РФФИ 16-04-00678.
Об авторах
Евгений Владиславович Даев
ФГБУ ВПО «Санкт-Петербургский государственный университет»
Автор, ответственный за переписку.
Email: mouse_gene@mail.ru
д-р биол. наук, профессор, кафедра генетики и биотехнологии Россия
Список литературы
- Tzapigina R, Aref’ev A, Sverdlova O, Daev E. Pheromonal regulation hypothesis of the space-genetic structure of the house mouse (Mus musculus L.) populations. World Congress of landscape ecology, IALE, Abstr. Ottawa, Canada; 1991. P. 84.
- Инге-Вечтомов С.Г. Экологическая генетика. Что это такое? // Соросовский образовательный журнал. — 1998. — № 2. — С. 59–65. [Inge-Vechtomov SG. Jekologicheskaja genetika. Chto jeto takoe? Sorosovskij obrazovatel’nyj zhurnal. 1998;(2):59-65. (In Russ).]
- Holttum S. Research watch: the power of genetic and biological explanations to reduce social inclusion. Mental Health and Social Inclusion. 2012;16(3):116-120. doi: 10.1108/20428301211255383/.
- LeVay S. Gay, straight, and the reason why: the science of sexual orientation. N.Y.: Oxford Univ. Press; 2011. P. 1-412.
- Definition of terms: sex, gender, gender identity, sexual orientation. In: The guidelines for psychological practice with lesbian, gay, and bisexual clients, adopted by the APA Council of Representatives; 2011. http://www.apa.org/pi/lgbt/resources/guidelines.aspx.
- American Heritage® Science Dictionary. Retrieved January 22, 2016 from Dictionary.com website http://dictionary.reference.com/browse/sexual-orientation.
- Agate RJ, Grisham W, Wade J, et al. Neural, not gonadal, origin of brain sex differences in a gynandromorphic finch. PNAS. 2003;100(8):4873-4878. doi: 10.1073/pnas.0636925100.
- Ngun TC, Ghahramani N, Sánchez FJ, et al. The genetics of sex differences in brain and behavior. Frontiers in Neuroendocrinology. 2011;31:227-246. doi: 10.1016/j.yfrne.2010.10.001.
- Ensemble genome browser release, 83. 2015. http://www.ensembl.org/Homo_sapiens/Location/Genome (посл. вход 02.09.2016).
- Cytochrome P450, family 21, subfamily A, polypeptide 2; CYP21A2. http://omim.org/entry/613815?search=cyp21&highlight=cyp21 (посл. вход 01.09.2016).
- Steroid 5-alpha-reductase 2; SRD5A2. http://omim.org/entry/607306?search=srd5a2&highlight=srd5a2 (посл. вход 01.09.2016).
- Cytochrome P450, family 19, subfamily A, polypeptide 1; CYP19A1. http://omim.org/entry/107910?search=cyp19a1&highlight=cyp19a1 (посл. вход 01.09.2016).
- Yamauchi Y, Riel JM, Ruthig VA, et al. Two genes substitute for the mouse Y chromosome for spermatogenesis and reproduction. Science. 2016;351(6272):514-516. doi: 10.1126/science.aad1795.
- Sanders AR, Martin ER, Beecham GW, et al. Genome-wide scan demonstrates significant linkage for male sexual orientation. Psychological Medicine. 2015;45:1379-1388. doi: 10.1017/S0033291714002451.
- Burri A, Cherkas L, Spector T, Rahman Q. Genetic and environmental influences on female sexual orientation, childhood gender typicality and adult gender identity. PLoS ONE. 2011;6(7):e21982. doi: 10.1371/journal.pone.0021982.
- Alexander GM, Wilcox T, Woods R. Sex differences in infants’ visual interest in toys. Arch Sex Behav. 2009;38:
- -433. doi: 10.1007/s10508–008–9430–1.
- Hines M. Prenatal endocrine influences on sexual orientation and on sexually differentiated childhood behavior. Frontiers in Neuroendocrinology. 2011;32:170-182. doi: 10.1016/j.yfrne.2011.02.006.
- McIntyre MH, Edwards CP. The early development of gender differences. Annual Review of Anthropology. 2009;38:83-97. doi: 10.1146/annurev-anthro-
- –164338.
- Ball GF, Balthazart J, McCarthy MM. Is it useful to view the brain as a secondary sexual characteristic? Neuroscience and Biobehavioral Reviews. 2014;46:628-638. doi: 10.1016/j.neubiorev.2014.08.009.
- Hines M, Brook C, Conway GS. Androgen and psychosexual development: Core gender identity, sexual orientation, and recalled childhood gender role behavior in women and men with congenital adrenal hyperplasia (CAH). The Journal of Sex Research. 2004;41(1):75-81. doi: 10.1080/00224490409552215.
- Kinsey, Alfred C, et al. Sexual Behavior in the Human Female. Philadelphia: W.B. Saunders; Bloomington, IN: Indiana U. Press; 1953.
- Beaulieu-Prévost D, Fortin M. The measurement of sexual orientation: Historical background and current practices. Sexologies. 2015;24:e15-e19. doi: 10.1016/j.sexol.2014.05.006.
- National Research Council. The Health of lesbian, gay, bisexualand transgender people: building a foundation for better under-standing. Washington, DC: The National Academies Press; 2011. From http://www.ncbi.nlm.nih.gov/books/NBK64806/ (посл. вход 01.09.2016).
- Andersen JP, Blosnich J. Disparities in Adverse Childhood Experiences among Sexual Minority and Heterosexual Adults: Results from a Multi-State Probability-Based Sample. PLoS ONE. 2013;8(1):e54691. doi: 10.1371/journal.pone.0054691.
- Corliss HL, Cochran SD, Mays VM. Reports of parental maltreatment during childhood in a United States population-based survey of homosexual, bisexual, and heterosexual adults. Child Abuse & Neglect. 2002;26:1165-1178.
- Plöderl M, Tremblay P. Mental health of sexual minorities. A systematic review. International Review of Psychiatry. 2015;27(5):367-385. doi: 10.3109/
- 2015.1083949.
- Muscarella F. The Evolution of Male-Male Sexual Behavior in Humans. Journal of Psychology & Human Sexuality. 2007;18(4):275-311. doi: 10.1300/J056v18n04_02.
- Edward DA. The description of mate choice. Behavioral Ecology. 2015;26(2):301-310. doi: 10.1093/beheco/aru142.
- Kohl JV. The Mind’s Eyes: Human Pheromones, Neuroscience, and Male Sexual Preferences. Journal of Psychology & Human Sexuality. 2006;18(4):313-369 (посл. вход 02.09.2016).doi: 10.1300/J056v18n04_03.
- Sela L, Sobel N. Human olfaction: a constant state of change-blindness. Exp Brain Res. 2010;205:13-29. doi: 10.1007/s00221–010–2348–6.
- Alexander BM, Skinner DC, Roselli CE. Wired on steroids: sexual differentiation of the brain and the role in the expression of sexual partner preferences. Frontiers in Еndocrinology. 2011;2:1-11. doi: 10.3389/fendo.2011.00042.
- McClintock MK. Estrous synchrony: modulation of ovarian cycle length by female pheromones. Physiol Behav. 1984;32:701-705.
- Russell MJ, Switz GM, Thompson K. Olfactory influences on the human menstrual cycle. Pharmacol Biochem Behav. 1980;13:737-738.
- Stern K, McClintock MK. Regulation of ovulation by human pheromones. Nature. 1998;392:177-179.
- Cadman SM, Kim S-H, Hu Y, et al. Molecular Pathogenesis of Kallmann’s Syndrome. Horm Res. 2007;67:231-242. doi: 10.1159/2F000098156.
- Garcia-Gonzalez D, Murcia-Belmonte V, Clemente D, de Castro F. Olfactory System and Demyelination. The Anatomical Record. 2013;296:1424-1434. doi: 10.1002/ar.22736.
- Martins Y, Preti G, Crabtree CR, et al. Preference for Human Body Odors Is Influenced by Gender and Sexual Orientation. Psychological Science. 2005;16:694-701. doi: 10.1111/j.1467–9280.2005.01598.x.
- Даев Е.В. Психогенетика с основами генетики. — СПб.: Н-Л, 2015. [Daev EV. Psihogenetika s osnovami genetiki. Saint Petersburg: N-L; 2015. (In Russ).]
- Stevenson RJ. An Initial Evaluation of the Functions of Human Olfaction. Chem Senses. 2010;35:3-20. doi: 10.1093/chemse/bjp083.
- Warr N, Greenfield A. The molecular and cellular basis of gonadal sex reversal in mice and humans. WIREs Dev Biol. 2012;1:559-577. doi: 10.1002/wdev.42.
- Berenbaum SA, Beltz AM. How early hormones shape gender development. Current Opinion in Behavioral Sciences. 2016;7:53-60. doi: 10.1016/j.cobeha.2015.11.011.
- Герлинская Л.А. Механизмы поддержания гетерогенного воспроизводства в популяциях млекопитающих: Автореф. дис. … д-ра биол. наук. — Новосибирск, 2008. [Gerlinskaja LA. Mehanizmy podderzhanija geterogennogo vosproizvodstva v populjacijah mlekopitajushhih. [dissertation]. Novosibirsk; 2008. (In Russ).]
- Kimball BA, Cohen AS, Gordon AR, et al. Brain Injury Alters Volatile Metabolome. Chemical Senses. 2016;00(00):1-8. doi: 10.1093/chemse/bjw014.
- Moshkin M, Gerlinskaya L, Morozova O, et al. Behaviour, chemosignals and endocrine functions in male mice infected with tick-borne encephalitis virus. Psychoneuroendocrinology. 2002;27:603-608.
- Lie HC, Simmons LW, Rhodes G. Genetic dissimilarity, genetic diversity, and mate preferences in humans. Evolution and Human Behavior. 2010;31:48-58. doi: 10.1016/j.evolhumbehav.2009.07.001.
Дополнительные файлы

